 当前位置:
当前位置:(本文来源:http://blog.sciencenet.cn/blog-1475614-807801.html)
首先,生态学发源于人类早期朦胧的生态意识。大约在数百万年前,据称一种非洲猿的站立(直立行走方式的进化是从猿类到原始人类的重要变革),无论是必然还是偶然,拉开了猿人→古智人→现代智人依次在自然历史舞台上粉墨登场的序幕。不管是“源自非洲”还是“多区域演化”,现代人类已经遍布与主宰了所有大陆。工具(石器等)的发明,逐渐推动了人类狩猎技巧的复杂化,随后农耕社会登场,迎来了人类自然生态意识的快速积聚。夸张地说,早期人类的发展史就是一部与人类生存技能相关的(或者说与应用性直接关联的)朴素生态意识的发展史。
朴素生态意识的科学化是人类对生命科学认识的积累与深化的产物。人类经历了对生命自身发展历程认识的曲折过程:1)中世纪西方特创论将世界万物描写成上帝的特殊创造物;2)从文艺复兴到18世纪,生物物种的不变论占据统治地位,这一时期,大量的生物物种被认识或描述,代表性人物—瑞典伟大的植物学家林奈(Carolus Linnaeus,1707-1778)建立了分类体系和双名制命名法;3)接着出现了“活力论”学说,代表性人物为法国著名的生物学家拉马克(Jean-Baptiste de Lamarck,1744-1829),虽承认物种可变,但归因于生物的“内部的力量”,认为这种非物质的活力驱动生物的进化,使之趋于复杂和完善;4)1858年,英国伟大的博物学家达尔文(Charles Robert Darwin,1809-1882)发表了著名的“物种起源”,他以非常“生态的”思想,揭示了生物进化的奥妙,即从生物与环境(包括生物的、非生物的)相互作用的视角,诠释了变异、遗传和自然选择如何推动物种进化。达尔文抛弃了上帝、不变和非物质的活力等朴素(或者说神秘的)生态意识,历史性地敲开了生命科学唯心论的一大缺口,使生态学得以诞生并从陈旧生命观的禁锢中脱缰而出,迎来了人类的科学生态思想的喷涌。本本主要是简述生态学(包括思想、重要概念等)早期的发展简史,解读生态学学科的分化,即寻根生态学。
一、生态学概念的起源与定义
任何一门科学发展到一定程度,都必须提出能最好地描述其特性的基本概念,生态学亦如此。概念—这一意识的载体,是人类对一个复杂事物或过程思辨和抽象后产生的,通过(可以是不同的)术语来表达。概念是一切旨在将本质与表象、必然与偶然区分开来的基石。那么,是谁最先提出了生态学的概念?
1. 生态学一词的起源
1859,进化论的创始人达尔文发表了著名的《物种起源》一书,这也是十九世纪生物学的最主要知识成就。七年后的1866年,著名的德国动物学家ErnstHaeckel在其专著“Generelle Morphologie der Organismen”(译:普通生物形态学)中首先使用了生态学(Oecologie)一词,Oikologie由两个希腊字oikos(house)和logos(study of)合并而成,从字面上来看,意指研究居住环境(或生境)的学科。当然,生态学还是属于生物学的一个分枝。
有学者认为生态学一词的发明权应归属于著名的美国博物学家Henry David Thoreau(他还是物候学之父),而另一些德国学者Schwarz和Jax(2011)认为Thoreau在此之前就使用了Oecologie一词则属于误传。
有学者分析,Haeckel发明Oecologie的初衷并不是要用他的概念、理论和实践来建立一个生态学学科,因为他自己决没有进行过所谓“生态的”研究。据推测,他可能只是想发明Oecologie一词来正身他的动物系统中当时尚未命名的一个分支学科而已,因为在该动物系统中,他用Oecologie来意指有机体的外部生理(externalphysiology of organisms);另一个证据就是,在这本著作中,他是先在一个示意图中使用了该词,然后才用文字予以解释(Jax and Schwarz 2011)。
很显然,Oecologie一词受到世界各国科学家的普遍认同,虽然经历了一段不短的时间。它于1869年传入俄国,出现于Haeckel(1866)的专著“Generelle Morphologie der Organismen”的俄文简译本中;于1874年传入法国,出现于1876年Haeckel(1868)的专著Natürliche Schöpfungsgeschichte的法译本。Oecologie的英文翻译似乎出现于1876年Haeckel专著的英译本TheHistory of Creation中,被翻译为oecology,后来被更改为ecology(Schwarz & Jax 2011)。1895年日本学者三好学(Miyoshi Manabu)将Ecology译为“生态学”,大约在1935年之前由武汉大学张挺教授传入中国,被广泛使用至今(阳含熙 1989)。
2. 生态学的定义
如果仔细留意一下,就不难发现几乎每本生态学教科书都会给生态学下一个定义,有些可能一样,有些可能类似或者说大同小异,而有些可能会明显不同。梳理一下这些生态学定义及其演变过程,也不是一件毫无意义的事。
Haeckel创作生态学一词的同时,也给它下了定义,虽然他在几个出版物(Haeckel 1866, 1868, 1870)中给Oekologie一词下了稍有不同的定义(Jax and Schwarz 2011),但是基本上是将生态学定义为研究生物与环境(包括生物的和非生物的)相互关系的科学,这可称之为“Haeckel式生态学定义”(表1)。
Haeckel(1866)最初将生态学看作“外部生理学(external physiology)”—“We divide physiology also into two disciplines: I. thephysiology of conservation or selfpreservation (a. nutrition, b. reproduction),II. the physiology of relations (a. physiology of the relations of parts of theorganism to each other (meaning, for animals, the physiology of nerves andmuscles); b. ecology and geography of the organism or physiology of therelations with the external world)”[译:“我们将生理学分为二个学科:I. 保持或自我维护的生理学(a.营养,b. 繁殖),II. 关系的生理学(a. 有机体的一部分与另一部分相互关系的生理学(指对动物来说,神经和肌肉的生理学);b. 有机体的生态学和生物地理学或与外界关系的生理学)”]。
早期的生态学定义基本上就是Haeckel式生态学定义,虽然这些定义在措辞上或多或少有些变化。这一概念无可非议,因为生物间以及生物与环境之间的相互作用确实是地球上所有生命系统(或称生态系统)的本质特性,它也是推动生物进化和生态系统演化的最基本驱动力。而一定空间中的生物与环境就构成了所谓的生态系统(这一概念稍后将要介绍)。但是对很多人来说,Haeckel式生态学定义有些过于抽象与宽泛,边界难以确定。
近半个世纪以来,虽然有一些学者仍然使用Haeckel的生态学定义,但也出现了一些新的生态学定义,体现在1)一些定义关注生物的分布和丰度,2)一些定义关注生态系统的不同层次—个体、种群和群落,3)一些定义关注生态系统的结构和功能。有一些则试图将好多种概念整合在一起(如Likens 1992),还有个别甚至包含了进化(如Petrides 1968)。
这种生态学概念的演化似乎反应了在过去的一个多世纪,人们的生态学认识在生命层次上从个体→种群→群落→生态系统的逐渐深化的过程,在时间尺度上从相对较短的响应向长时期的动态变化、从相对简单的要素向复杂生态系统的结构与功能解析的转变过程。应该说,所有这些生态学概念都反应了生态学涵括的不同侧面,也都是真实的。不得不承认,由于生态系统类型、过程、格局及尺度等呈现出的极端的多样性与复杂性,定义一个普遍接受的生态学概念,可能除了抽象和宽泛,确实也很难有别的选择。
表1关于生态学的代表性定义
Table 1 Representative definitions of ecology
|
生态学的定义(英文) Definition of ecology (in English) |
译文In Chinese |
文献References |
|
By ecology we mean the whole science of the relations of the organism to its surrounding outside world, which we may consider in a broader sense to mean all ‘conditions of existence’. These are partly of an organic nature and partly of an inorganic nature. |
生态学指有机体与外部世界的环境之间相互关系的所有科学,这在广义上指生存条件,一部分是有机性质的,另一部分是无机性质的 |
|
|
The ecology of the organisms, the science of the whole relations of organisms to their surrounding world, towards the organic and inorganic conditions of existence; the so-called ‘economy of nature’, the interrelations of all organisms which live in one and the same place, their adaptations to their environment, their transformation through the struggle for existence |
有机体的生态学,即有机体与其周边世界的所有关系的科学,包括有机和无机的生存条件;所谓‘自然的经济学’,即生活在一个同样地方的所有有机体的相互关系,它们对环境的适应性,以及通过生存斗争的变化 |
Haeckel 1868 |
|
By ecology, we mean the science of the economy, of the household of animal organisms. This has to study the entirety of relations of the animal both to its inorganic and its organic environment, in particular the benign and hostile relations with those plants and animals with which it comes directly into contact; or, to be concise, all those intricate interrelations which Darwin calls the struggle for existence |
生态学指研究动物居住环境经济学的科学。这不得不研究动物与无机和有机环境之间的所有关系,特别是与与之直接接触的那些动植物之间的有益和有害的关系;或者,简单地说,所有那些达尔文称之为生存斗争的?相互关系 |
Haeckel 1870 |
|
the scientific natural history concerned with the sociology and economics of animals |
与动物的社会学和经济学有关的科学自然历史 |
Elton 1927 |
|
the science of all the relations of all organisms to all their environments |
所有生物与它们的所有环境所发生的所有关系的科学 |
Taylor 1936 |
|
|
|
|
|
the science of the inter-relation between living organisms and their environments, including both the physical and biotic environments, and emphasizing interspecies as well as intraspecies relations |
生物与环境之间相互作用的科学,包括物理和生物环境,强调种间和种内关系 |
Allee et al. 1949 |
|
In its broadest sense, the science of ecology can be defined as the study of the relations between plants and animals and their environment; it will then include most of biology, biochemistry and biophysics. In its narrower sense, ecology is taken to refer to the study of plant and animal communities |
广义地说,生态学可定义为研究植物和动物之间及其与环境之间的相互关系,它将包括生物学、生物化学和生物物理学的大部分内容,狭义地说,生态学指关于植物和动物群落的研究 |
Clarke 1954 |
|
the science which investigates organisms in relation to their environment: a philosophy in which the world of life is interpreted in terms of natural processes |
研究生物与其环境相互关系的科学,一种生物界用自然过程来诠释的思想体系 |
Woodburry 1954 |
|
a science which concerns itself with the inter-relationships of living organisms, plants and animals, and their environment |
与生物体(植物和动物)及其环境内在关系相关的科学 |
|
|
the scientific study of the distribution and abundance of organisms |
研究生物分布和丰度的科学 |
|
|
the study of animals and plants in relation to each other and to their environment |
研究动物和植物之间及其与环境之间关系的科学 |
Kendeigh 1961, 1974 |
|
the study of interactions of form, functions and factors.’ |
研究类型、功能和因子相互作用的科学 |
Misra 1967 |
|
the study of the way in which individual organisms, populations of some species and communities of populations respond to these changes |
研究个体、一些物种的种群和种群形成的群落对其变化响应方式的科学 |
Lewis and Taylor 1967 |
|
the study of environmental interactions which control the welfare of living things, regulating their distribution, abundance, production and evolution |
研究控制生物的福利、调控其分布、丰度、生产及进化的环境相互作用的科学 |
Petrides 1968 |
|
Margalef defined ecology as the biology of ecosystems |
Margalef将生态学定义为生态系统的生物学 |
Margalef 1968 |
|
the study of the structure and function of ecosystems or broadly of nature |
研究生态系统(或广义的自然)的结构或功能的科学的科学 |
Odum 1971 |
|
the study of ecosystems, or the totality of the reciprocal interactions between living organisms and their physical surroundings |
研究生态系统、生物与其物理环境之间所有相互作用的科学 |
Clark 1973 |
|
the study of relations between organisms and the totality of the biological and physical factors affecting them or influenced by them |
研究生物与其影响和被影响的所有生物环境、物理环境相互关系的科学 |
Pinaka 1974a |
|
the scientific study of the relationships of living organisms with each other and with their environments |
研究生物之间及与环境之间相互关系的科学 |
Southwick 1976 |
|
a multidisciplinary science which deals with the organisms and its place to live and which focuses on the ecosystem |
关于生物和生境的多学科的科学,聚焦生态系统 |
Smith 1977 |
|
the scientific study of the interactions that determine the distribution and abundance of organisms |
Krebs将生态学定义为研究决定生物分布和丰度的相互作用的科学 |
Krebs 1978 |
|
The scientific study of the processes influencing the distribution and abundance of organisms, the interactions among organisms, and the interaction between organisms and the transformation and flux of energy and matter |
研究影响生物分布和丰度的过程、生物之间的相互作用、以及生物与能量和物质转换和流动之间相互作用的科学 |
Likens 1992 |
二、生态学早期的发展简史
有学者(Egerton1977, McIntosh 1985)认为,Haeckel创造了生态学一词,但这并不意味他就创造了生态学,因为Oecologie一词的出现并没有立刻激发起大量的生态学研究,在其后的20多年的时间里,很少有人注意到这一新术语,甚至他本人也未能进行有效的生态学研究;也就是说,Haeckel为生态学提供了一个名称,却几乎没有给这门学科提供实质性内容。
那么,生态学到底源自何处?这或许是一个像定义生态学一样难于回答的问题,而且不同的学者也有很不相同的观点,但是,有一点是毫无疑问的—大多数学者还是认为生态意识或思想源远流长。
1. 生态学的起源—从本能到科学
Voorhees(1983)专门创造了一个有趣的名称—protoecologist(前生态学家)来意指那些在生态学成为一门正式的科学之前具有生态学见解的人们。正是由于生态学的过于宽泛的定义,导致了关于“哪些人是前生态学家”、“谁是生态学之父”或“谁是第一个生态学家”或“谁创建了生态学”这样的问题十分难于回答,一直争论不休,甚至带有一些民族主义色彩。有学者甚至极端地宣称“自古以来每个植物学家都是生态学家”(Greene 1909)。即便如此,刻意回避这一问题,就无法翻开生态学发展历史上的精彩一页。其实,科学的生态学思想也是人类及人类社会意识进步的产物。
1)本能的生态感觉
由于生态学指生物与环境(生物、非生物的)之间的相互关系,而所有的生物都必须“懂得”生态的关系才能得以生存下来,譬如一头初生的老虎必须在成长的过程中从母虎那儿学习各种狩猎的技巧、并不断的摸索和实践,才得以成功捕获猎物并生存下去,这是一种直觉的(或本能的)生态意识,没有它,物种(动物)不可能存在。同样的道理,最原始的人类如果没有丰富的与各种生物的习性相关的生态学知识,就不可能成功地进行渔猎活动。可以这样说,自然选择塑造了各种动物在一定环境中生存的本能的生态感觉或意识。
2)朦胧或朴素的生态意识
从猿人→古智人→现代智人(自然界最高级的动物)的人类进化历程与其对自然生态知识的不断积聚密不可分。人类文明的重要象征就是文字的发明,其必然结果就是人类运用文字对身边的动植物(包括命名)及相关自然生态现象的记述,随着生态知识的不断积累,必然会导致一些朦胧或朴素的生态思想的形成。
事实上,如果按照Haeckel的原始生态学定义,即研究生物与环境(生物、非生物的)之间的相互关系,那历史上一切与之关联的动植物生活习性的描述(如散落在各种史书—农书、药物书、专谱、地方调查记、杂记等之中)等都可以算作生态学。
早在2000多年前,世界古代史上最伟大的哲学家、科学家—古希腊的亚里士多德(公元前384—322年),就研究和描述了500多种动物,他是将生物分门别类的第一人,并将一些生态因素(如栖居地、生活习性、生活方式等)也整合进动物分类中,他详细地观察、记录或思考了一些动物的生态习性(如鱼类的捕食行为、鱼卵的损失情况等),并撰写了相关的著作如《动物志》、《动物的迁移》、《动物的运动》等。这些著作,特别是有关动物栖息地的描述,无不明确地印记着朴素的生态学思想,只是没有贴上时髦的“生态学”的标签而已。
我国最早的词典—《尔雅》,距今也有2000多年的历史,它由19篇组成,其中多达7篇(即释草、释木、释虫、释鱼、释鸟、释兽、释畜)是关于动植物方面的词语,对许多动植物的形态和生活环境进行了描述或记载,也是我国古老而朴素的生态思想的汇集。
3)科学或认知的生态思想
所谓科学是指在理性、客观的前提下,通过一套必要的方法,对自然作理由充分的观察或研究,获得有组织体系的知识,揭示自然真相或证明自然真理。因此,科学的生态学思想与朦胧或朴素的生态思想的最大不同应该在于前者发展成了生态学的知识体系。
科学生态学的诞生是博物学在生命科学领域发展的必然产物
可以这样说,科学生态学(scientific ecology)就是博物学的知识结晶。所谓博物学是指对大自然的宏观观察和分类,也即所谓“自然史”(natural history)研究,虽然后来随着科学的分化与深入,博物学逐渐衰落,但在科学与文明史中博物学的贡献功不可没,十八~二十世纪初期是伟大的博物学家辈出的时代,如林奈、拉马克、达尔文等。
博物学的诞生可以称得上是人类对自然(包括生态)现象认识上的一次重要飞跃,它使人类开始运用较为系统的科学方法来观察与认识自然,特别是使人类对自然生态的认识从局域的生活环境走向地理区域乃至全球,极大地推动了人类综合思维的发展,在生命科学方面的硕果就是诞生了一系列重要的关于生命与生态现象的规律性的认识(如进化论)。可以这样说,博物学也是现代科学技术飞速发展的前奏。
有理由相信,只有像博物学家对生态现象进行系统的归纳与综合并抽象出规律,才会使人类对生态的认识进入科学生态学时代。事实上,一些早期的生态学家把传统的博物学视为科学生态学的开端,而达尔文的进化论又主要是博物学的产物,因此包括Haeckel(一个达尔文主义的强烈拥护者)在内的许多早期生态学家将科学生态学归功于达尔文及其提出的由自然选择引起的进化论(进化生物学)思想也就不足为奇了(McIntosh 1985)。
有意思的是,Jax和Schwarz(2011)认为Haeckel并未意识到博物学对生态学起源的重要意义,因为Haeckel(1870)宣称生态学构成了“自然史”的主要部分:“Ecology (often also inappropriately calledbiology in the narrower sense) has, up to now, constituted the main componentof so-called ‘natural history’ in the usual sense of this word.”。
著名的英国动物生态学家Elton(1927)在其经典之作“Animal Ecology”中将生态学称为“科学的自然史”(the scientific natural history concernedwith the sociology and economics of animals)不正是绝妙地影射出自然史与科学生态学之间的这种血脉关系吗?
达尔文是“藐视”生态学一词的科学生态学的奠基人
如果翻开达尔文的《物种起源》(Darwin 1859)一书,能看到无数关于种间与种内竞争以及基于竞争的自然选择等生态学现象的描述、分析和推理,当然他关注的核心是物种的起源。譬如达尔文认为“全世界所有生物之间的生存斗争,这是它们按几何级数高度增值的不可避免的结果”,“每一个物种所产生的个体,远远超过其可能生存的个体,因而便反复引起生存斗争,于是任何生物所发生的变异,无论多么微小,只要在复杂而时常变化的生活条件下以任何方式有利于自身,就会有较好的生存机会,这样便被自然选择了”,“同种的个体间和变种间的生存斗争最为激烈,同属的物种间的斗争也往往激烈”,等等。再看看Haeckel的生态学定义,将达尔文的进化论(进化和适应)视为科学生态学之父应该不是子虚乌有。
有趣的是,达尔文并未在意他对生态学的贡献,因为他甚至在生态学一词被创造(1866年)之后也从未使用过该词,甚至在这之后他的很“生态”的著作如于1881年发表的“通过蚯蚓的作用形成腐殖土”(The formation of vegetable mould throughthe action of worms)中也未使用过。这似乎是说,达尔文进行了真正的开拓性生态学研究,却没有在生态学的名义下进行!
同样有趣的是,那个时代的一些生态学奠基者们在其工作中也几乎没有涉及达尔文的思想(Acot 1997),可能主要是因为启蒙时期的生态学主要关注有机体形态和生理与非生物环境之间的相互关系,即所谓的环境选择性,而达尔文的自然选择主要聚焦于生物间的竞争作用(Paterson 2005)。
当然,也有一些学者并不认同达尔文是生态学之父的说法,譬如,Gendron(1961)主张著名的博物学家Alexander von Humboldt(1767–1835)开创了生态学,因为他通过植被的地理学研究,提出了植被垂直分布的思想;甚至有学者认为,公元前4世纪的古希腊哲学家和科学家—Theophrastus才称得上历史上第一个生态学家,因为他明确地记述了植物聚集在一起形成群落,以及植物之间以及植物与无生命环境之间的关系(Ramaley1940)。
2.达尔文的进化论—马尔萨斯人口论学说在整个动植物界的应用
1831年,年仅22岁的达尔文随英国海军的勘探船“小猎犬号”进行了为期五年的环球考察,对大量的地质地理现象、生物化石、形形色色的动植物物种的形态与生态等进行了详细的考察、纪录与分析,正是在这次漫长的环球航行中积累的丰富的博物学知识,使他萌生了生物学史上最重要的一个观念,也是一个关于人类自身在自然界位置的伟大的生命观:即人类绝非上帝制造,只不过是生命进化的产物罢了(齐默2011)。
有意思的是,被视为科学生态学之根的达尔文的进化生物学的思想却从人口论的研究(即关于人类种群数量变动的研究)中获得了很大启发。大约在《物种起源》(Darwin 1859)问世前的半个多世纪,一个名叫马尔萨斯(Thomas Malthus)的英国乡村牧师于1798年发表了《人口学原理》(An Essay on the Principle of Population),这是一本以悲观论调讨论人类却难的书籍,其核心观点是:①人口呈几何级数(即:2,4,8,16,32,64,128等)增长;②而食物供应呈现算术级数(即:1,2,3,4,5,6,7等)增长;③因此,必定存在限制人口过度增长的机制,这就是贫困与罪恶,表现为失业、疾病、饥荒、瘟疫、暴行、战争等等。也即马尔萨斯强化了资源的有限增长与人口的无限增长之间的矛盾以及所导致的残酷的“生存挣扎”的必然后果。人口呈几何级数、食物供应呈现算术级数增长可用图1来形象地表示。对达尔文来说,种群过度繁殖与资源有限性之间的矛盾不可避免地导致大部分子代个体提前夭折是自然选择的前提条件,因为如果每个个体都能存活到生理极限的话,就不存在择优了。
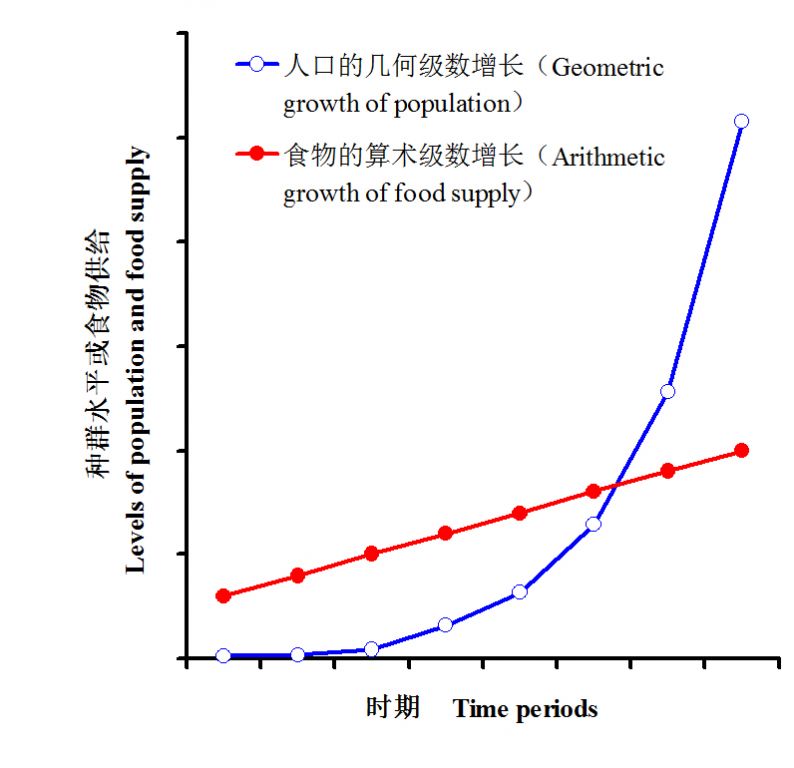
图1 马尔萨斯提出的人口呈几何级数增加、食物呈算数技术增加示意图
指数增长的威力可借用道金斯(1981)的一段话来予以说明:“拉丁美洲目前的人口大约有三亿,而且其中已有许多人营养不良。但如果人口仍按目前的速度继续增长,要不了五百年的时间,人口增长的结果就会出现这样一种情况:人们站着挤在一起,可以形成一条遮盖该大陆全部地区的由人体构成的地毯。即使我们假定他们都是瘦骨嶙峋—一个不是不真实的假定——情况依然如此。从现在算起,在一千年之后,他们要相互立在肩膀上,其高度要超出一百万人。待到两千年时,这座由人堆起的山将会以光速向上伸展,达到已知宇宙的边缘”。这看起来太骇人听闻,但却隐喻了指数增长的威力!
资源是否是等差数列增长(在一定范围内也许是等比数列增长)其实已不很重要,但地球资源的有限性这一点是毫无疑问的。马尔萨斯还指出,多产与饥荒这两种控制人口的力量,同样也在控制动植物,即如果苍蝇生蛆完全不受障碍,世界很快将蛆满为患,因此,大部分苍蝇必须在未能繁殖后代前死去(齐默2011)。
正是马尔萨斯的这一“无限”与“有限”之间的矛盾,对达尔文产生了关键性的影响,后者吸取了生存斗争的基本思想,发展了自然选择的概念:一方面,有机体会出现或大或小的变异或突变(现代的科学技术已经证实这种突变积累在一种称之为DNA的遗传物质上),这种变异可能会改变个体的生存能力;另一方面,在自然界,动植物个体可能会为了光、水、食物、空间等资源而竞争(种间的或种内的)。因此,物种就是这样经历着一种没有筛选者的筛选,正是这种无需任何创造动作干预的筛选,在漫长的历史岁月中塑造了各式各样看似“精密设计”的生命。换言之,达尔文从马尔萨斯的悲观的人口论中找到了推动生命进化的动力(齐默2011)。
达尔文在其经典著作—《物种起源》(Darwin 1859)中也承认,他的理论是“马尔萨斯学说在整个动物界和植物界的应用”。从这种意义上说,达尔文的进化论似乎也应该是马尔萨斯的生态学思想的产物之一。
3.马尔萨斯人口论的数学描述—逻辑斯蒂方程式
二十世纪20年代以前,生态学在静态的描述性研究上停滞不前,大量的论文局限于描述某一区域的物种数量及个体,而研究种群数量变动的种群生态学的登场使这一局面得到了扭转。
其实,早在19世纪30年代,数学家就对马尔萨斯的人口论产生了兴趣,并提出了描述人口变动的著名的逻辑斯蒂模型。遗憾的是,这一模型沉寂了相当长的时间,直到1920年才被重新关注,但是几年后,在此基础上Lottka和Voltera提出了著名的描述捕食者-猎物相互作用的数学方程式,种群生态学(Population ecology)从此步入了发展的黄金时期,引发了大量的生态学家去关注与定量地描述种群的行为,如生长、衰退、周期性波动等。那么,什么是逻辑斯蒂模型?
1)无限增长模型—指数增长
在理想(不受食物、空间等限制)的情况下,一切生物物种均呈现一种无限增长的模式(也即所谓与密度无关的增长density independent growth)。
在理想的条件下生物物种的种群增长模式可用下式描述:
dN/dt = rN (1.1)
积分后得到:
N(t) = N(0) exp(rt) (1.2)
这里N为种群密度,r为常数,是种群在没有密度制约因素条件下的增长速率的度量,r也常被称为“马尔萨斯参数Malthusian parameter”或“内禀增长率intrinsic growth rate”。式1.2可以看成是马尔萨斯关于人口在理想情况下无限增长思想的数学描述。
从式1.2不难看出,若r>0,则种群无限增长;若r<0,则种群指数式下降。事实上,正如马尔萨斯指出的那样,人口不可能无限增长,更广泛地说,在自然界由于食物、空间等因子的限制,没有哪一个物种(无论是动物还是植物)能无限增长的,最多在一定的时期内能呈现这种特征(如近代人类的人口增长)。也即,现实世界中的任何物种都终究会呈现一种与密度有关的增长(density dependent growth)模式。其实,种群增长是环境有限性与物种增殖无限性之间均衡的结果。那么,如何用数学方程式来描述像马尔萨斯叙述的那种种群的有限增长?
2)有限增长模型—逻辑斯蒂增长
19世纪早期人们对人口增长的数学理论的兴趣明显增加,其代表性人物就是著名的比利时数学家Pierre François Verhulst。在马尔萨斯的人口论发表40年之后,Verhulst(1838)用下述数学模型来描述了马尔萨斯的人口理论:
dN/dt=rN(1-N/K) (1.3)
这里,N为种群(人口)密度,r为内禀增长率(intrinsic growth rate),不受环境制约;K为环境容量(carrying capacity),它决定于食物、空间或其它;每个个体的增长率与密度有关,即r(1-N/K),若N<K,为正值,若N>K,为负值;而在N=K时,种群达到平衡。
将式(1.3)积分后,得到:
N(t)=K/(1+CKe-rt) (1.4)
这里,N(t)为时间t时的个体数, C = 1/N(0)−1/K。
Verhulst在其1845年发表的论文中称式(1.4)为逻辑斯蒂函数(logistic function),现在称为逻辑斯蒂方程式(logistic equation)。有学者也称之为Malthus-Verhulst逻辑斯蒂理论(Berryman 1992)。
有意思的是,逻辑斯蒂模型居然发表在达尔文的“物种起源”(1859年)之前,但是,这一模型一直未能被人关注,直到1920年,Raymond Pearl和LowellReed重新发现了这一模型(图2)。
人类—是地球上最有影响力的动物种群,人口的增长对地球环境正在产生前所未有的巨大影响。我们还看不到人类的种群何时能呈现出一个漂亮的逻辑斯蒂曲线,也许我们还要等数个世纪或更长,但终究它会达到增长的极限。
自1866年Haeckel创造生态学一词以来,生态学沉寂了接近半个世纪,发展十分缓慢,到了20世纪20年代才得到广泛承认,并发展成一门生机勃勃的科学,那时,也开始了理论数学种群生态学研究的黄金时期。到了20世纪60年代,“环境危机”突然将生态学置身于社会公众的舞台,并被广泛推崇,甚至成了政治口号(McIntosh 1985)。
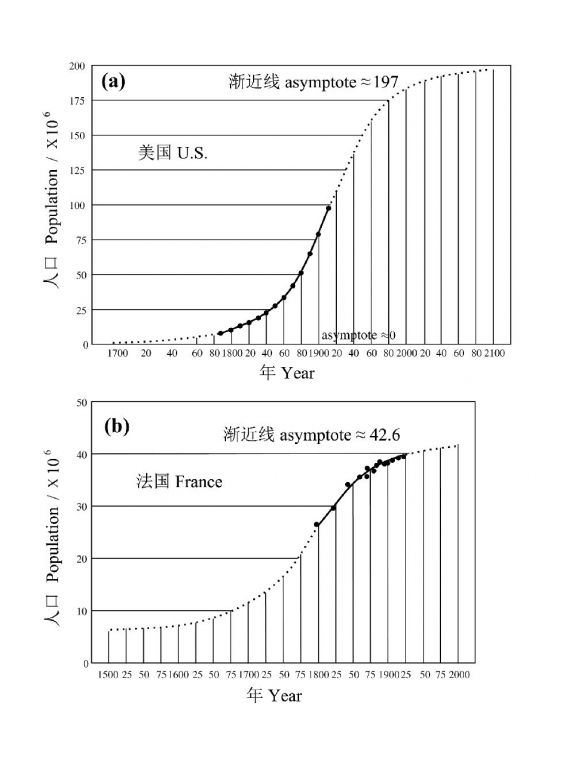
图2用于拟合美国(a)和法国(b)人口数据的逻辑斯蒂种群增长(式1.4)。决定参数的数据仅涵盖了增长曲线的一小部分(根据Pearl 1925)(引自Murray2002)
三、生态学早期的其它重要概念
1.生物圈(Biosphere)
1875年,奥地利地质学家Eduard Suess创造了biosphere一词,定义为“地球表面生物居住的地方”,是地球的四大圈层之一,即大气圈(atmosphere)、水圈(hydrosphere)、岩石圈(lithosphere)和生物圈(biosphere)。二十世纪20年代,生物圈一词获得了其生态学意义(Huggett 1999),代表性人物是俄国地球化学家Vladimir Vernadsky,他进一步发展了生物圈的概念,认为生物与行星环境(planetary environment)协同进化并形成了生物生存的介质(空气、水、土壤、沉积物),其生物圈观点最初于1926年用俄文发表(Vernadsky1926)。生物圈的范围一般指海平面以上约10000 m至海平面以下10000 m处,包括大气圈的下层,岩石圈的上层,整个土壤圈和水圈,但绝大部分生物集中在地表以上100 m到水下100 m的大气圈、水圈、岩石圈、土壤圈等圈层的交界处,也是生物圈的核心。从空间尺度上来看,生物圈虽然是地球表面薄薄的一层,但仍然是一个浩瀚的生态系统。
2. 生物群落(biocenosis)
1877年,德国动物学家Karl Möbius创造了biocenosis一词(Möbius 1877),描述生活在一个生境(habitat)中相互作用的有机体,认为每一个生物群落都“支持着一定数量的有生命的生物”,强调在合适的条件下可能产生过量的后代,但由于空间和食物是有限的,“群落中个体总数不久又会回到它以前的适中状态”,这使人回想起马尔萨斯关于人口与它们受资源限制的理论(McIntosh 1985)。但在英语中biocenosis一词很少使用,而常用Ecological community,指占据同一地理区域的二个或以上不同种类的物种的集合。
3. 生物地理学(biogeography)
1891年,德国地理学家Friedrich Ratzel创造了biogeography一词(Parenti & Ebach 2009),而德国博物学家Alexander von Humboldt(1769-1859)被誉为植物地理学的创始人(von Humboldt 1805),他走遍了西欧、北亚和南、北美洲进行了植物学和地质学考察,揭示了植物分布的水平和垂直分异性及其与气候的关系,发现植物形态随高度而变化,并将植物在世界范围内进行分区,确立了植物区系的概念。美国生态学家Robert MacArthur和Edward Osborne Wilson创建了岛屿生物地理学理论,揭示了一定面积的物种多样性能通过生境面积、移居速度、灭绝速率等进行预测(MacArthur & Wilson 1967)。
4. 生态位(niche)
1917年,美国动物学家Joseph Grinnell创造了Niche一词(Grinnell 1917),意指某一物种的生态位是由其居住的生境所决定的,而Elton(1927)的生态位观念强调物种在群落中的功能,而不是生境;1957年,英裔美国动物学家George Evelyn Hutchinson提出了n维超体积(n-dimensional hypervolume)的生态位空间的概念(Hutchinson 1957),试图解释为何在同一个生境中有如此之多的不同类型的生物共存在一起。
5.食物链(food chain)
1927年,英国动物生态学家Charles Elton在其经典之作“AnimalEcology”中引入了food chain的概念,他借用中国谚语(Chineseproverbs)来诠释food chain的含义:“Thelarge fish eat the small fish; the small fish eat the water insects; the waterinsects eat plants and mud”(译:大鱼吃小鱼、小鱼吃昆虫、昆虫吃植物和泥巴)。食物链概念的重要生态学意义在于揭示了物质和能量在物种之间的转移与流动。与此相联系,Elton还借用中国的谚语“One hill cannot shelter two tigers”(译:一山不容二虎)诠释了Pyramid of numbers(数量金字塔)的概念,即越在食物链顶端(即营养级越高),动物的数目越少。Elton还开拓性地提出了foodcycle的概念(在后来的文章中用food web取代了foodcycle)。在Elton数量金字塔的基础上,美国生态学家Raymond L. Lindeman提出了能量转换的生态效率(ecological efficiency of energy transfer)的概念,即能量从低营养级到高营养级的转化效率(Lindeman 1942)。
6. 生态系统(Ecosystem)
1930年,英国植物学家Roy Clapham创造了Ecosystem一词(Willis 1997),由英国生态学家Tansley(1935)在印刷的论文(题为The use and abuse of vegetational conceptsand terms)中正式使用,并被详细地诠释,旨在强调一个既包含了生物又包含了环境的系统,即认为生物有机体虽然是这些系统中的最重要的组成,但是无机因素也是组成部分,相互作用不仅发生有机体之间,也发生在有机与无机成分之间。
大约在1905年前后,美国著名的植物生态学家Clements提出了关于植物群落演替的所谓“super-organisms(超级有机体)”概念(McIntosh 1985)。Clements(1916)认为顶极群落构成的就是一个超有机体,因为呈现出与单株植物十分相似的生命历程—产生,发展,成熟,死亡...。Tansley(1935)是在讨论这一“超级有机体”概念时引入了“生态系统”一词,这也是一场在Clements与Tansly之间展开的关于植物界是一个超级有机体(super-organism)还是一个生态系统的争论。
形象地说,Tansley是把“生态学”装在一个称之为“系统”的盒子里了,再冠以“生态系统”的名称!但是他所定义的这个生态系统有时清楚,有时又是十分模糊,因为他赋予了它最为多种多样的类型和大小。虽然Tansley认为生态系统是自然界的基本单元,也得到了广泛的认可,但常常是生态系统的边界无法明确界定,这在现代生态学依然是一个经常困扰着很多人的难题。
与生态学一词类似,生态系统一词从其被创造到被广泛应用于生态科学,其间也有一段时间的间隔(McIntosh 1985)。
世界上没有哪种物质形态能像生命这样如此多姿多彩。不论是在非生命还是生命学科,可能也没有哪一个学科能像生态学这样如此多样。借用植物学的比喻来说,它似一颗枝叶繁茂的大树,与其它学科以及各种生物类群、各种生境等组合成了各式各样的生态学。
1. 生态学的分类
因为生态学泛指一切关于生物与环境相互关系的科学,在这种宽泛而抽象的生态学定义下,就免不了出现各式各样的生态学分枝(表1-2)。譬如,以生态系统的层次为基础,分化出分子生态学、个体生态学、种群生态学、群落生态学、生态系统生态学、景观生态学、全球生态学等;生态学也经常以生物对象来命名:植物生态学(以植物类群甚至某个种细分)、动物生态学(以动物类群甚至某个种细分)、微生物生态学等等;生态学还以生物栖居的环境为基础,划分为水域生态学(再细分为:海洋生态学、湖泊生态学、河流生态学……)和陆地生态学(再细分为:森林生态学、草原生态学、荒漠生态学、土壤生态学、城市生态学……)等等;生态学与各种学科进行交叉,分化出生理生态学、化学生态学、进化生态学、古生态学、数学生态学等等;生态学还与产业交叉,形成农业生态学和工业生态学等;生态学还被广泛移植到人文社科领域,产生出社会生态学、政治生态学、文化生态学等等,甚至被移植到医学领域,出现药物生态学、健康生态学等等。生态学家还以组合或叠加的方式创作出各式各样的生态学名称。
表1-1生态学的分类
|
生态学名称 |
外文生态专著举例 |
中文生态专著举例 |
|
1. 生命层次 |
|
|
|
分子生态学Molecular ecology |
Freeland 2005 |
祖元刚等1999 |
|
种群生态学Population ecology |
Begon et al. 1996 |
徐汝梅1987(注:昆虫种群生态学) |
|
Tilman & Kareiva 1997 |
— |
|
|
集合种群生态学Metapopulation ecology |
Hanski 1999 |
— |
|
群落生态学Community ecology |
Diamond & Case 1986 |
赵志模和郭依泉 1990 |
|
植被生态学Vegetation ecology |
van der Maarel 2009 |
姜恕和陈昌笃1994 |
|
系统生态学System ecology |
Odum 1983 |
蔡晓明2000 |
|
流域生态学Watershed ecology |
Naiman 1992 |
— |
|
景观生态学Landscape ecology |
Forman & Godron 1986 |
傅伯杰2011 |
|
全球生态学Global ecology |
Rambler et al. 1989 |
方精云2000 |
|
2. 学科交叉 |
|
|
|
生理生态学Physiological ecology |
Townsend & Calow 1981 |
蒋高明 2004(注:植物生理生态学) |
|
营养生态学Nutritional ecology |
Slansky & Rodriguez 1987 |
— |
|
营养(级)生态学Trophic ecology |
Mbabazi 2011 |
— |
|
代谢生态学Metabolic Ecology |
Sibly et al. 2012 |
— |
|
生物物理生态学Biophysical Ecology |
Gates 1980 |
— |
|
化学生态学Chemical ecology |
Sondheimer & Simeone 1970 |
阎凤鸣 2003 |
|
进化生态学Evolutional ecology |
Pianka 1978 |
王崇云2008 |
|
地理生态学Geographical ecology |
MacArthur 1972 |
— |
|
地生态学Geoecology |
Huggett 1995 |
— |
|
古生态学Paleoecology |
Dodd & Stanton 1981 |
杨式溥1993 |
|
第四纪生态学Quaternary ecology |
Delcourt & Delcourt 1991 |
刘鸿雁 2002 |
|
环境生态学Environmental Ecology |
Freedman 1989 |
金岗等1992 |
|
污染生态学Pollution ecology |
Hart & Fuller 1974 |
王焕校1990 |
|
水文生态学Hydro-ecology |
Wood et al. 2007 |
— |
|
历史生态学Historical ecology |
Crumley 1994 |
— |
|
稳定同位素生态学Stable isotope ecology |
Fry 2006 |
易现峰2007 |
|
理论生态学Theoretical ecology |
May 1976 |
张大勇2000. |
|
数学生态学Mathematical ecology |
Pielou 1977 |
陈兰荪1988 |
|
数字生态学Numerical ecology |
Legendre & Legendre 1998 |
— |
|
数量生态学Quantitative ecology |
Poole 1974 |
张金屯2004 |
|
统计生态学Statistical ecology |
Young & Young 1998 |
— |
|
实验生态学Experimental ecology |
Resetarits & Bernardo 2001 |
— |
|
3. 生物类别 |
|
|
|
植物生态学Plant ecology |
Warming 1895 |
张玉庭和董爽秋1930 |
|
作物生态学Crop ecology |
Loomis & Connor 1992 |
韩湘玲1991 |
|
动物生态学Animal ecology |
Elton 1927 |
费鸿年1937 |
|
昆虫生态学 Insect ecology |
Speight et al. 1999 |
邹钟琳1980 |
|
鸟类生态学Avain (bird) ecology |
Perrins & Birkhead 1983 |
高玮1993 |
|
鱼类生态学Fish ecology |
Wootton 1992 |
易伯鲁1980 |
|
渔业生态学Fisheries ecology |
Pitcher & Hart 1982 |
陈大刚1991(注:黄渤海渔业生态学) |
|
野生生物(动物)生态学Wildlife ecology |
Moen 1973 |
陈化鹏和高中信 1992 |
|
杂草生态学Weed ecology |
Radosevich & Holt 1984 |
— |
|
寄生虫生态学Parasite ecology |
Huffman & Chapman 2009 |
— |
|
微生物生态学Microbial ecology |
Alexander 1971 |
夏淑芬和张甲耀. 1988 |
|
疾病生态学Disease ecology |
Learmonth 1988 |
— |
|
4. 生境类型 |
|
|
|
森林生态学Forest ecology |
Spurr & Barnes 1973 |
张明如2006 |
|
草地生态学Grassland ecology |
Spedding 1971 |
周寿荣. 1996 |
|
海洋生态学Marine ecology |
Levinton 1982 |
李冠国和范振刚. 2011. |
|
河口生态学Estuarine ecology |
Day et al. 1989 |
陆健健2003 |
|
潮间带生态学Intertidal ecology |
Raffaelli & Hawkins 1996 |
— |
|
海岸生态学Coastal ecology |
Barbour et al. 1974 |
— |
|
淡水生态学Freshwater ecology |
Macan 1974 |
何志辉2000 |
|
湖泊生态学Lake ecology |
Scheffer 2004 |
— |
|
河流生态学River ecology |
Whitton 1975 |
— |
|
溪流生态学Stream ecology |
Allan 1995 |
— |
|
湿地生态学Wetland ecology |
Keddy 2010 |
陆健健等2006 |
|
水库生态学Reservior ecology |
Tundisi & Straškraba 1999 |
韩博平等2006 |
|
城市生态学Urban ecology |
Bornkamm et al. 1982 |
于志熙 1992 |
|
道路生态学Road ecology |
Forman 2003 |
— |
|
廊道生态学Corridor ecology |
Hilty et al. 2006 |
— |
|
土壤生态学Soil ecology |
Killham 1994 |
曹志平2007 |
|
5. 动植物行为与功能 |
|
|
|
行为生态学Behavioral ecology |
Krebs & Davies 1997 |
尚玉昌1998 |
|
扩散生态学Dispersal ecology |
Bullock et al. 2002 |
— |
|
繁殖生态学Reproductive ecology |
Bawa et al. 1990 |
张大勇2004 |
|
摄食生态学Feeding Ecology |
Gerking 1994 |
— |
|
认知生态学Cognitive Ecology |
Friedman & Carterette 1996 |
— |
|
功能生态学Functional ecology |
Packham et al. 1992 |
— |
|
6. 环境扰动与胁迫 |
|
|
|
扰动生态学Disturbance ecology |
Johnson & Miyanishi 2007 |
— |
|
火生态学Fire ecology |
Wright & Bailey 1982 |
— |
|
胁迫生态学Stress ecology |
Steinberg 2011 |
— |
|
7. 产业与应用 |
|
|
|
工业生态学Industrial ecology |
Graedel & Allenby 2002 |
邓南圣和吴峰2002 |
|
农业生态学Agricultural ecology |
Azzi 1956 |
曹志强和邵生恩 1996 |
|
资源生态学Resource ecology |
Prins & van Langevelde 2008 |
— |
|
恢复生态学Restoration ecology |
Jordan III et al. 1990 |
赵晓英和陈怀顺 2001 |
|
应用生态学Applying (or Applied) ecology |
Beeby 1993 |
何方2003 |
|
8. 组合或叠加 |
|
|
|
传粉与花的生态学Pollination and floral Ecology |
Willmer 2011 |
— |
|
陆地植物生态学Terrestrial plant ecology |
Barbour et al. 1989 or 1999 |
— |
|
理论系统生态学Theoretical ecosystem ecology |
Ågren & Bosatta 1998 |
— |
|
微生物分子生态学Molecular microbial ecology |
Osborn & Smith 2005 |
张素琴2005 |
|
鸟类迁移生态学The migration ecology of birds |
Newton 2008 |
— |
|
应用数学生态学Applied mathematical ecology |
Levin et al. 1989 |
— |
|
应用野外生态学Practical field ecology |
McLean & Ivimey Cook 1946 |
— |
|
数量植物生态学Quantitative Plant Ecology |
Greig-Smith 1957 |
— |
|
10. 人文社会与人体健康 |
|
|
|
深生态学Deep Ecology |
Devall & Sessions 1985 |
雷毅2001 |
|
人类生态学Human ecology |
Hawley 1950 |
陈敏豪1988 |
|
社会生态学Social ecology |
Alihan 1964 |
丁鸿富1987 |
|
人口生态学Population ecology |
Davis 1971 |
潘纪一1988 |
|
政治生态学Political ecology |
Cockburn & Ridgeway 1979 |
刘京希2007 |
|
组织生态学Organizational ecology |
Hannan & Freeman 1989 |
刘桦2008 |
|
文化生态学Cultural ecology |
Netting 1986 |
邓先瑞和邹尚辉. 2005 |
|
嵌套生态学Nested ecology |
Wimberley 2009 |
— |
|
道教生态学Toaism ecology |
|
乐爱国2005 |
|
Mühlhäusler 1996 |
— |
|
|
健康生态学Health ecology |
Hunarī et al. 1999 |
— |
|
药物生态学Pharma-ecology |
Jjemba 2008. |
— |
2.为何有如此繁多的生态学分支出现?
为何出现如此繁多的生态学也不是一件特别容易回答的问题。从根本上来看,生态(包括生物类群、生境类型、生存环境、生命过程、生命演化等)的复杂性可能主导性地决定了生态学科的多样性。譬如:
1)生态学要面对一个庞大而变化多样的生物类群:地球上现存的生物物种超过170万,小的种类的个体不足1微米,大的可达150多米(植物)或190吨(动物),种类极为纷繁,且跨越巨大的生命(体积)尺度;
2)生态学要涉及一系列空间跨度极为巨大的生态系统:小可到一个烧杯大可到整个生物圈;
3)生态学要涉及一系列事件跨度极为巨大的生态过程:短可仅为数分钟,长可涉及数十亿年的生物演化;
4)生态学要面对的生物的生存条件跨越巨大的气候梯度:从寒冷的极地冰川,到炎热的热带区域,年平均降雨量从0.5 mm(南美洲智利共和国最北端的阿里卡)到超过12000 mm(印度的乞拉朋奇),等等。
5)生态学要面对的生物生存的垂直梯度从海拔-416 m的地表(死海)到海拔超过8000 m的高山(珠穆朗玛峰),从水陆交接的海岸带到超过11000 m的深海(马里亚纳海沟),跨越巨大的物理化学等环境梯度;
6)生态学要涉及各种各样地貌特征完全不同的生境,如湖泊、河流、水库、湿地、森林、草地、农田、海洋等等,以及这些生境之间异常复杂的交融与相互作用,等等。
简言之,可能没有哪一类学科像生态学这样,试图在相当精细的程度上,面对如此繁多的研究对象和生境类型,跨越如此宽广的时空尺度,包含如此之大的气候与环境梯度以及如此之多样的地貌类型。生态学的多样性从本质上来看正是其所关注对象(物种、群落、生态系统、格局、过程等)多样性的一种映射。
这里,引用普里戈金和斯唐热(1987)的一句名言:“科学通过把现实的复杂性约化为一种隐藏的简单性而得到进步”,这也是笔者的核心目的,即试图从复杂纷繁、恢宏壮阔的生命现象中凝析出生命系统的生态学(当然还有与之相适应的遗传学)设计原理,揭示我们人类赖以生存的地球系统中精彩绝伦的生命系统存在与演进的秘诀。
不可否认,人类正在日益肆无忌惮地任意地摆弄与操纵(或者说近乎疯狂地统治与奴役)着自然界及其进程,正在近乎随心所欲地不断改造与创造“新的大自然”。因此,我们需要认知生命世界的本质、组织方式以及置生于其中的人类的位置与角色。而且,我们还需要更多的生态智慧去提升认识自身的活动对未来影响的预见能力,尽可能地去填充在人的利欲设计与大自然的生态设计之间存在的鸿沟。
五、结语
生态学的发展经历了从本能的感觉(渔猎时代)、朦胧的意识(早期人类文明)到科学的体系(现代科学时代),从起源上看,科学生态学是博物学发展的产物。由于对象的多样性,衍生出种类繁多的生态学分枝。虽然早期的科学生态学思想可追溯自达尔文的进化生物学研究(物种起源),之后生态学脱离进化生物学踏上了相对独立的发展之道。
参考:从生态学透视生命系统的设计、运作与演化—生态、遗传和进化通过生殖的融合(谢平著,科学出版社,2013)
联系方式:沈宏 徐军 027-86780056 Email:wetland@ihb.ac.cn
地址:武汉市武昌区东湖南路7号
访问数量:
