 当前位置:
当前位置:(本文来源:http://blog.sciencenet.cn/blog-1475614-815185.html)
如果你将踏上环球之旅,你或许会去北美的索罗兰沙漠触摸神奇的仙人掌,或去南美体验亚马逊热带雨林的壮观,或去中国西部的内蒙古眺望辽阔的大草原,或去中国北部的黑龙江欣赏独特的三江湿地风光(图1)。此外,如果你环绕热带,则可以观赏到完全不同的自然景象—干枯荒芜的沙漠中难觅生命的踪迹,稀树点缀的大草原上狮子—角马呼啸奔腾、叶子退化的刺林显露出干渴而粗壮的枝干,神秘幽深的热带雨林中满是“树上生树”、“叶上长草”的奇妙景观!
令人称叹的是,植物学家用了一个非常贴切的词语来形容这些各具特色的植物群落—“植被”,它似乎是分类学和生态学的“混血儿”,既易于被专业的植物学家所接受,也易于被非植物学人士乃至普通大众所理解。
毫无疑问,这些覆盖在地表的植物群落(即植被)是地球上一切其它生命存在的基础,亦为漫长的地质历史长河中生命演化的产物。当然,植被地理格局具有一定的南北对称性,虽然不完全对称。那么,为何不同的地理区域会出现完全不同的植被类型?一个不容置疑的事实是,地球上许多独特的植被类型与不同的气候带遥相呼应,折射出二者之间若隐若现或千丝万缕的联系!从古至今的农林牧业生产实践与近代大量的植物生理生态学研究已经证实,温度、光照、降雨和养分是决定植物生长的最基本环境要素,而且这些要素密切联系在一起(譬如太阳光照决定了温度的基本格局),其中温度和降雨被认为在决定植被地理格局中最为重要。

图1 世界各地不同植被类型的比较:(A)美国亚利桑那仙人掌国家公园中的仙人掌(图片引自jdnx),(B)巴西亚马逊热带雨林(图片引自Destination360),(C)中国西部的内蒙古大草原(图片引自ChinaTour.Net),(D)中国北部黑龙江的三江湿地(图片引自大树)
一、如何描述与分类自然界的植物集群?
为了描述植物特征的方便,人们先后创造了不同的名词,本节将简要地介绍植物群落、生活型、植被、生物群系、植物区系等概念。植物群落的空间尺度如何?这也是本节需要探讨的问题。
1. 常见的概念—植物群落、生活型、植被、群系和区系
植物群落(plant community,偶尔也用phytocoenosis或phytocenosis)指在特定空间中生活在一起的各种植物种类的集合,植物群落的命名也常多样而灵活,如苔藓群落、漂浮植物群落、热带雨林、温带草原等。植物群落的特征主要由种类组成、丰度、外貌和水平—垂直结构等来描述。
生活型(life-form):指适应于一定生镜具有相似外貌特征的植物群落类型,常用来描述高等植物。这种外貌特征可以指高矮、大小、形状、分枝等,对有些植物类群还结合植物的年度周期(一年生或多年生)来区分。譬如,可将高等植物分为木本植物(乔木、灌木、藤本)和草本植物(多年生草本、一年生草本),木本植物还可分为阔叶和针叶,以及落叶和常绿等类型;也可以生境为特征将高等植物分为陆生植物、水生植物、湿生植物等类型。可以这样说,生活型是综合考虑植物形态学、分类学和生态学等的一种混合植物群落分类方法,重点强调植物的外貌和环境适应性。
实际上,系统发育上密切相关的植物可能有完全不同的生活型,如菊科植物有高不足 5厘米的一年生小草本,也有高达10米的热带或亚热带乔木。相反,无亲缘关系的种类也可能通过趋同(convergence)进化出同样的生活型。
植被(vegetation)是地表覆盖的植物的总称。植被可由单一植物群落或多个植物群落组成,如有些植被主要由森林群落组成,而另一些植被则由森林、灌丛和草甸等群落组成。
生物群系(biome)是指基于优势生物类群(如植物、动物、微生物)和气候类型所划分的地理尺度的生物群落。常常以植被类型作为特色,故又可指地带性植被。全球尺度看即所谓植物群系地球系统。
植物区系(flora)指某一特定地区生长的全部植物种类,通常将某地区全部植物种类按科、属、种进行数量统计,然后按地理分布、起源地、迁移路线、历史成分和生态成分划分成若干类群,分别称为植物区系的地理成分、发生成分、迁移成分、历史成分、生态成分等。动物区系则成为fauna。植物区系、动物区系和所有其它生命形式(如真菌)一起统称为生物区系(biota)。
植物群落、生活型、植被、群系和植物区系等概念既有区别,又有联系,不同学者也有不同的解读,有些概念有时也被混用。一般来说,植被比植物区系的概念要宽泛,后者仅指物种组成;植被与植物群落的意思相同,但常常涉及更大的空间尺度(包括全球)。
2. 空间尺度—多大才算一个植物群落?
人们经常谈论植物群落,但是,到底起码多大才算一个植物群落?而不同植物的个体差异极大,如小的植物—浮游藻类直径仅有1-2 μm,而树木—澳洲的杏仁桉最高可达156 m。因此,可被认知的特定植物群落的空间尺度也应该差别巨大。
在野外研究中,植物生态学家为了研究一定植物群落的生态现象,需确定合适的样方面积。表1是van der Maarel(2005)综合了数位学者的看法给出的各种植物群落的最小面积,最小的植物群落面积只需不足1m2(附者苔藓和地衣群落),最大的可达10000 m2(热带雨林)。
表1各种植物群落的最小面积(m2)
|
植物群落类型 Types of plant communities |
|
面积Area (m2) |
|
附者苔藓和地衣群落Epiphytic moss and lichen communities |
|
0.1–0.4 |
|
陆地苔藓和地衣群落 Terrestrial moss and lichen communities |
|
1–2 |
|
漂浮水生植物群落Free-floating aquatic communities (Lemnetea) |
|
2–5 |
|
湿生先锋群落 Hygrophilous pioneer communities (Isoeto-Nanojuncetea) |
|
2–5 |
|
踩踏栖息地的植被Vegetation of trampled habitats (Polygono-Poetea annuae) |
|
2–5 |
|
低位盐沼泽Lower salt marshes (Thero-Salicornietea) |
|
4–10 |
|
开放沙丘和沙质草地 Open dune and sand grasslands (Koelerio-Corynephoretea) |
|
4–10 |
|
(亚)地中海一年生植物群落 (Sub-)Mediterranean therophyte communities (Helianthemetea guttati) |
|
4–10 |
|
高强度经营的草场Heavily managed grasslands (Cynosurion cristati) |
|
4–10 |
|
高位盐沼泽Upper salt marshes ( Juncetea maritimi) |
|
10–25 |
|
根生漂浮水生植物Rooted floating aquatic communities (Potametea) |
|
10–25 |
|
温带草场Temperate pastures and meadows (Molinio-Arrhenatheretea) |
|
10–25 |
|
好盐土草场Basiphilous grasslands (Festuco-Brometea) |
|
10–25 |
|
雨养型沼泽植被Ombrotrophic bog vegetation (Oxycocco-Sphagnetea) |
|
10–25 |
|
泥潭和泥沼植被Bog-pool and mire vegetation (Scheuchzerio-Caricetea fuscae) |
|
10–25 |
|
干草原Steppes (Festuco-Brometea) |
|
20–50 |
|
(亚)高山钙质草原(Sub-)Alpine calcareous grasslands (Elyno-Seslerietea) |
|
20–50 |
|
海岸黄沙丘群落Coastal yellow dune communities (Ammophiletea) |
|
20–50 |
|
高沼泽植被Tall swamp vegetation (Phragmito-Magnocaricetea) |
|
20–50 |
|
石南灌丛Heathlands (Calluno-Ulicetea) |
|
20–50 |
|
杂草群落Weed communities (Stellarietea mediae) |
|
40–100 |
|
长廊林和林隙植被Woodland fringe and gap vegetation |
|
40–100 |
|
多年生杂草植被Perennial ruderal vegetation (Artemisietea vulgaris) |
|
40–100 |
|
温带灌木Temperate scrub (Rhamno-Prunetea) |
|
40–100 |
|
地中海灌木林Mediterranean maquis (Quercetea ilicis), chaparral |
|
40–100 |
|
地中海矮灌木Mediterranean low scrub (Cisto-Lavanduletea) |
|
40–100 |
|
柳树和杨树灌木林Willow and poplar scrub and woodland (Salicetea purpureae) |
|
100–250 |
|
高山硬叶灌木群落 Fynbos |
|
100–250 |
|
欧洲肥沃土壤上的落叶林 Deciduous forest on rich soils in Europe (Querco-Fagetea): |
草本层 herb layer |
100–250 |
|
沼泽丛林Swamp woodland (Alnetea glutinosae) |
|
100–250 |
|
针叶林 Coniferous forest (Vaccinio-Piceetea) |
|
200–500 |
|
欧洲肥沃土壤上的经营林Managed deciduous forest on rich soils in Europe (Querco-Fagetea): |
冠层canopy |
200–500 |
|
欧洲肥沃土壤上的成熟落叶林Mature deciduous forest on rich soils in Europe (Querco-Fagetea): |
冠层canopy |
400–1000 |
|
如上,在北美Ibid. in North America: |
冠层canopy |
400–1000 |
|
荒漠植被Desert vegetation |
|
400–1000 |
|
(亚)热带干旱林(Sub-)Tropical dry forest |
冠层canopy |
400–1000 |
|
热带次生林Tropical secondary forest |
冠层canopy |
2000–5000 |
|
热带雨林Tropical rain forest |
冠层canopy |
4000–10000 |
3. 主要植被类型—基于优势植物生长型的划分
植物学家对植被的分类研究已有很长的历史。早在十九世纪初,Humboldt就提出了根据植物群落的外貌来区分植被类型的分类系统(表2),这些植被类型反映了与气候类型密切关联的优势植物的生长型。
这样的分类系统强调植物群落的整体外貌,而不是拘泥于严格的植物分类系统,因此对即便是不太熟悉植物分类学的人来说也比较容易理解,因此被学术界(甚至社会大众)广泛接受和使用。
表2 主要植被类型及其特点
|
植被类型 Types of vegetation |
主要特点 Main characteristics |
|
1. 森林Forest |
高8米以上 |
|
(1)常绿林(针叶、阔叶) |
|
|
(2)落叶林(针叶、阔叶) |
|
|
2. 林地woodland |
2-8米的小高位芽植物 |
|
3. 灌丛Scrub |
低于2米的木本植物 |
|
4. 草地Grassland
|
草本(通常是禾草或苔草)是优势种,木本植物或缺如,或矮态而不显著 |
|
5. 稀树干草原Savannah |
|
|
(1)灌木稀树干草原Shrub savannah
|
矮高位芽植物,个别散布于稠密的草本植物、地衣的被盖之上 |
|
(2)树丛Groveland |
除最高层的植物聚成小树丛外,与savannah相似 |
|
6. 疏树草原Parkland
|
树丛的交互连接伴随有较低层草被,茂密地遍布于森林或林地的连续相中 |
|
7. 草甸Meadow
|
稠密草地,非禾草,伴生有叶片相当宽阔而柔软的禾草类,生境湿润; |
|
8. 干草原Steppe |
在高地上,在对森林来说过于干燥的区域的草地 |
|
(1)草甸性草原Meadow-steppe
|
干草原区域干燥程度低的边缘,类似草甸,低矮灌木常见,但不占优势 |
|
(2)真草原True steppe
|
区系贫乏和相当旱生的干草原,禾草叶片狭窄,并有贫乏的非禾草如灌木; |
|
(3)灌丛干草原Shrub-steppe
|
散生灌木突出于草本以上的草原; |
|
9. 草本沼泽Marsh
|
潮湿或周期性潮湿的,拥有矿质土壤的草地 |
|
10. 木本沼泽Swamp
|
潮湿或周期性潮湿的,拥有矿质土壤的木本植被 |
|
11. 荒原Fellfield |
冻原内不连续的低矮植被,地上芽植物最显著,土壤高度石质; |
(修改自曲仲湘等1984)
4. 主要植物群系—地带性植被类型
迄今为止,虽然如何界定生物群系的大小或依照什么标准来分类生物群系仍存争议,但这并不影响人们对这一概念的使用。世界主要陆地植物群系被分为若干类型,包括热带雨林、温带阔叶林、极地冻原等(表3),这些群系本质上反映了植被与地域、气候特征的强烈耦合。
表3世界主要陆地植物群系类型
|
热带雨林(包括高山云雾林)Tropical rain forest (including montane and cloud forests) |
|
热带落叶林、林地和多刺灌丛Tropical deciduous forest, woodland and thorn-scrub |
|
热带稀树大草原Tropical savanna |
|
沙漠Deserts |
|
暖沙漠(亚热带)Warm desert (subtropical) |
|
冷冬荒漠(大陆性的)Cold-winter desert (continental) |
|
温带森林Temperate forests |
|
落叶阔叶林Deciduous broad-leaved forest |
|
常绿阔叶林(包括月桂林、暖温带混交林)Evergreen broad-leaved forest (incl. laurel forest, warm-temperate mixed forest) |
|
温带雨林Temperate rain forest |
|
草原(温带)Grasslands (temperate) |
|
针叶林Conifer forests |
|
北方的(包括落叶的)Boreal (including deciduous) |
|
山地针叶林(温带山地和亚高山)Montane conifer forest (temperate montane and subalpine) |
|
冻原高山植被Tundra and alpine vegetation |
|
极地和温带—高山冻原Polar and temperate-alpine tundra |
|
热带高山植被Tropical alpine vegetation |
|
陆地湿地(木本沼泽、草本沼泽、泥炭沼泽、沼池)Terrestrial wetlands (swamp, marsh, bog, fen) |
(引自 Boxand Fujiwara 2005)
二、植被的地理格局—全球气候系统的产物
从全球看植被的分布,即植被的地理格局。植物地理学家很早就认识到气候与植被地理分布之间的密切关系,可以这样说,植被的地理格局就是地球上气候格局的产物。
1.全球的气候系统如何划分?
Walter(1984,1985)提出的全球气候类型系统(图2)可能是最被广泛接受的系统,它重视气候的核心区域而不是边界。而全球的气候带格局又是由全球大气环流及其季节移动形成的。全球大气环流系统的季节性南北移动使热带辐合带、亚热带高压带和其它组分周年性地影响一些纬度,而仅季节性地影响另一些纬度。许多主要的气候类型是地带性能量输入和全球环流的直接结果,而中纬度的一些类型也在很大程度上受到其在陆块上的地理位置的影响。
当然需要指出的是,看似对称的南北半球,实际也存在一定程度的非对称性:南半球的陆地面积要比北半球小得多,因此偏冷及更具海洋性气候特性,温度赤道(temperature equator)大约位于地理赤道以北10o(沃尔特1984)。
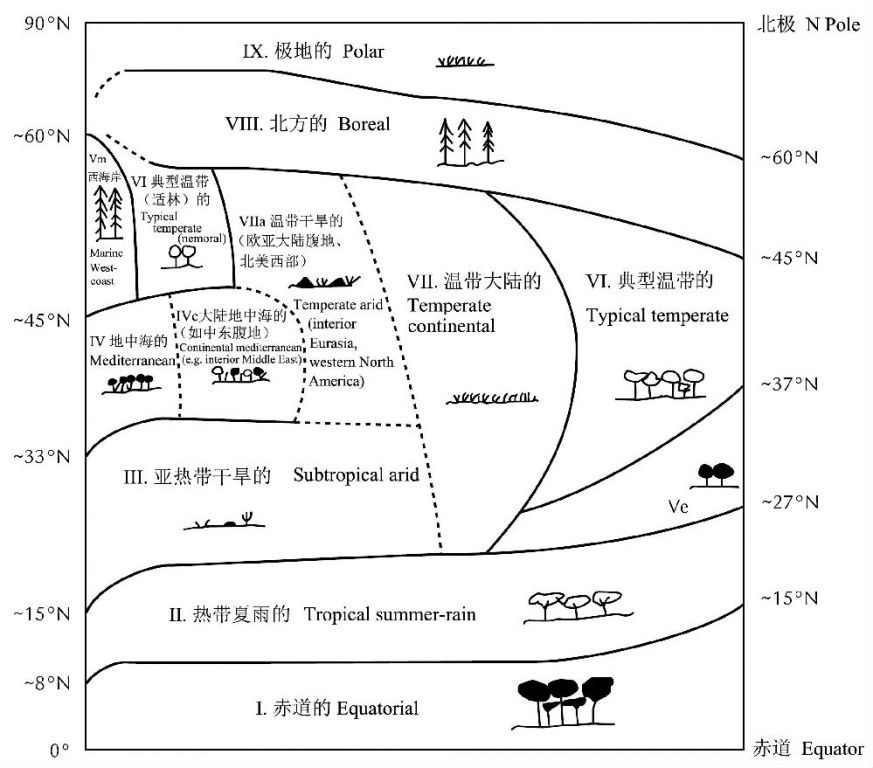
图2 一个理想大陆的气候区域(引自Box and Fujiwara 2005)
2.气候系统与地带性植被的耦合
Walter气候系统很好地反应了不同气候情势下特定植物类型的潜在发育机制,因为它很好地对应了世界主要的生物群系及其位置(表5-4)。
譬如,赤道气候(气候I)地带的植被为热带雨林,发生在无持续干旱期的地方,且温度决不会降低到10 oC以下,植物能周年生长等。在热带夏季雨林气候区(II),由于有明显的干湿季,适应于不同的年降雨范围,形成了三个不同的带状植被类型:在湿端(湿季长于旱季)的热带湿润落叶林、在中间的干燥落叶林和林地(如在非洲中南部的坦桑利亚林地)和在干燥端的雨绿多刺灌丛或稀疏大草原。
表5-4全球气候系统类型与地带性植被的对应关系
|
Climatic conditions |
Walter类型 Walter type |
地带性植被类型 Zonal vegetation types |
|
赤道的Equatorial |
I |
热带雨林(暖、湿) Tropical rain forest (warm, wet) |
|
湿/干(热带)Wet/dry (tropical) |
II |
雨绿林和林地(加上:热带稀树草原和多刺灌丛) Rain-green forests and woodlands (plus savannas and thorn-scrub) |
|
干旱(热、干) Arid (hot, dry) |
III |
暖半荒漠和荒漠 Warm semi-deserts and deserts |
|
地中海的(干夏) Mediterranean (dry summer) |
IV |
Sclerophyll forests and shrublands |
|
西海岸(冷、海洋的) Marine west coast(cool, oceanic) |
Vm |
温带雨林 Temperate rain forests |
|
暖东海岸Warm east coast |
Ve |
常绿阔叶林 Evergreen broad-leaved forests |
|
温带大陆性的Temperate continental |
VI |
Summer green forests |
|
VII |
温带草原Temperate grasslands |
|
|
VIIa |
温带半沙漠和沙漠 Temperate semi-deserts and deserts |
|
|
北方的Boreal |
VIII |
北方林Boreal forests |
|
极地的Polar |
IX |
冻原和冷荒漠Tundra and cold deserts |
(引自Box and Fujiwara 2005)
3.二个主要气候因子—温度和降雨对地带性植被的塑造
温度和降雨是二个最重要的气候因子(当然,对植物的生长和分布来说,光照和营养也是重要影响环境因子),强烈地影响着植被的地理格局。图3描绘了植物群系分布与气候因子之间的密切关系。
在湿润和炎热的气候条件下,常绿阔叶雨林占据优势,当气候向干、冷转化时,落叶林占据优势,而当气候趋于极端寒冷或干燥的状况下,优势植被被冻原或沙漠植被所取代。
同样是热带地区,随降雨量的逐渐增大,植被从叶子退化的刺林、稀树干草原变化到树木高大、物种丰富、生长密集且常绿的热带雨林。
在较低的降雨条件下,随着温度从低到高,植被从冻原(由矮灌木、苔藓、地衣、禾草和苔草等为主)→北方草地→温带草原→刺林逐渐变化。
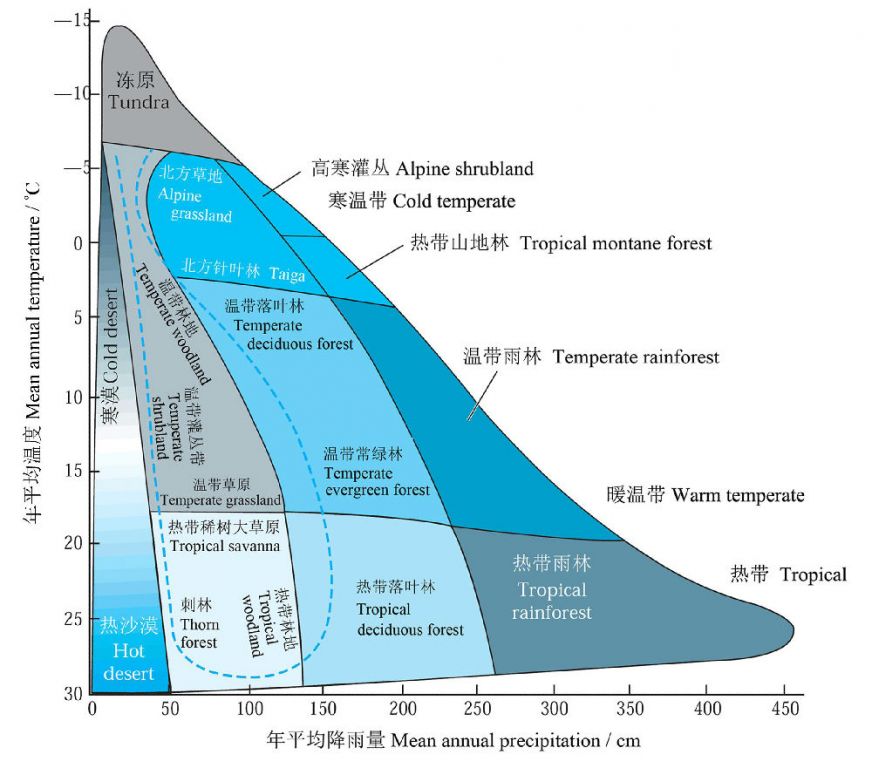
图3 植物群系分布与气候因子(年平均温度和降雨量)之间的关系。在虚线区域,其它因素(如火烧、牧食和降雨的季节性)强烈地影响群系类型。气候也能与其它因子(如土壤类型)相互作用决定植物群系的分布(引自Gurevitch et al. 2002)
有学者统计了世界范围内主要的植被类型(热带雨林、稀树干草原、温带落叶林、北方针叶林和冻原)在不同的月平均最低温度和总年降雨量条件下的分布情况(如图4所示),也能看出最低温度和降雨量对植被类型的明显影响,即热带雨林主要分布在高温多雨区域(虽然跨越很宽的降雨量范围),而冻原和北方针叶林则主要分布在低温干燥区,稀疏干草原分布于干热气候区,而温带落叶林分布于中等程度的水热条件区。

图4 陆生植物经历的气候条件可用年降雨量和平均月最低温度表示,(A)热带雨林,(B)稀树干草原,(C)温带落叶林,(D)北方针叶林(针叶林带)和(E)冻原(Begon et al. 2006 仿Heal etal. 1993)
很显然,极端的低温和高温对植被地理格局的影响绝然不同,在极端低温的情况下,生命受到极度的压抑,只有物种稀少、群落单一的耐寒性强的冻原得以存在。而在极端高温的热带地区,降雨的分配主宰了植被的地理格局,由于形成了极大的水分梯度,植被类型变化巨大:从极为干旱、物种极为贫瘠的热沙漠→刺林→热带落叶林→多雨的、物种极为丰富的热带雨林。高温、强蒸发和活跃的生命活力是热带地区植被演化的驱动机制。
4.局域环境因子的差异也能显著改变植被类型
即便是同样的区域气候背景,很多陆地生态系统呈现出相当多样的地貌变化,如在一些山区。因此,同一个区域,局域地形(地貌)造成的小气候条件或土壤质地等的差异,往往能能显著影响植物群落的结构。譬如,同样的石质山原峡谷,朝北面主要生长着喜冷、喜荫和喜湿环境的植物,而向南面生长着喜欢温暖、阳光和干燥环境的植物(图5,Gibson & Gibson 2006)。

图5 一个石质山原峡谷的朝北(左)和朝南(右)面。朝北面适合喜冷、喜荫和喜湿环境的植物生长(蓝叶云杉、花旗松和山枫),向南面适合于喜欢温暖、阳光和干燥环境植物的生长(西黄松、丝兰、仙人掌)(引自Gibson & Gibson 2006)
水陆交错带是一类受水位影响很大的生态系统,水位变化模式决定着水生植物、湿生植物与陆生植物群落分布的相对格局及动态。譬如,水位波动幅度能对沿岸带湿地植被的带状分布格局产生显著影响(图6),较大的水位波动导致湿地植被出现更为多样的带状格局,而当这种波动减少后,植被类型减少,导致群落的物种多样性下降(Cronk & Fennessy 2001)。
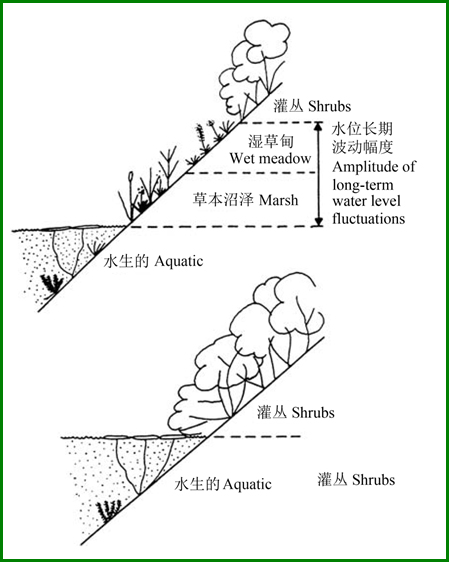
图6 稳定水位使湿地植被带从4种压缩至2种的概念示意图(引自Keddy 2000)
植被不仅收到地表水的影响,还受到地下水的影响。譬如,在美国的威斯康辛州,在同样的区域,地下水流量和排水状况不同引起的水文条件差异可导致多种多样的植被类型,即便所处的景观地貌类似(图7)。当然,在许多情况下,地表水和地下水是密切相关的。
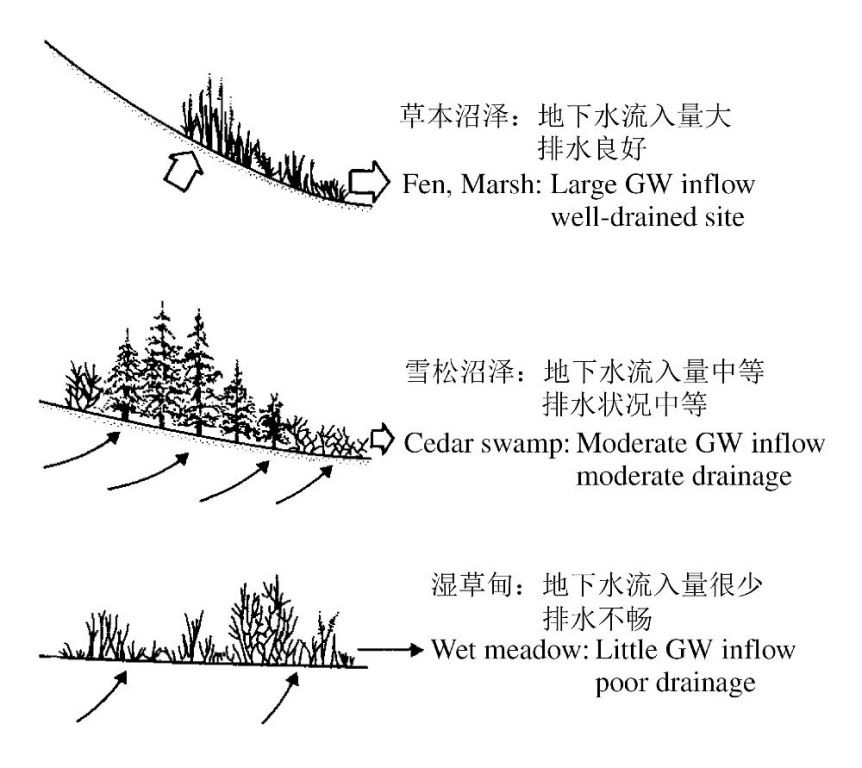
图7 美国威斯康辛州水文条件对湿地植被类型的影响(van der Valk 2006重绘自Novitzki 1979)
5. 沿海拔高度出现山地植被类型的明显更替
在同样的地带性气候背景下,植被沿山地的垂直方向呈现出显著不同的分布带,即海拔高度也能具有与纬度类似的对气候与植被的影响。从地貌上来看,山地沿垂直方向一般可分略为低丘陵、山地、高山和雪带等(沃尔特1984)。
气温沿纬度和海拔高度均表现出梯度性分布,即温度从赤道向极地逐渐降低,也随海拔高度上升而下降:从北向南每增加100 km或海拔每上升100 m温度增加1oC,这种现象被称为直减率(lapserate)。
植物学家观察到随海拔和随纬度梯度变化过程中具有十分类似的植物群系变化趋势(图8),当然,一般来说,山地植被高度带将比平原植被带狭窄约1000倍,但是二者也不会是简单的复现,因为在一些方面(如温度和光照节律、降雨量、土壤质地等)还是存在一定的差异。
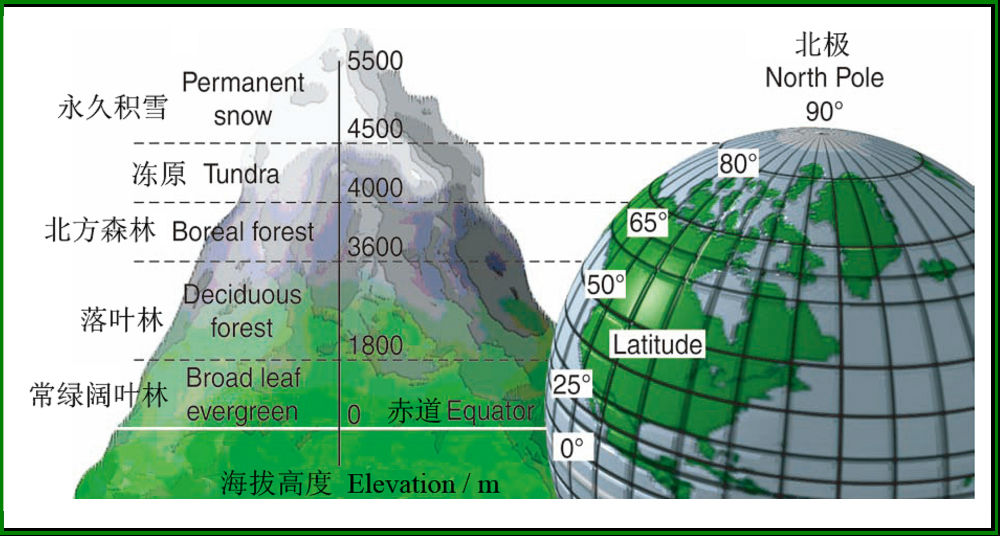
图8 海拔高度和纬度对气候和生物群系的分布具有相似的影响(引自Gibson & Gibson 2006)
如果调查沿海拔梯度上的各种植物的分布范围,再换算成温度,人们就可以粗略地推测各种树生存的温度范围。如在欧洲阿尔卑斯山上,各个树种分布的温度范围如图9所示。当然,人们可以推测这些树种的分布明显受到温度的控制,也许的确如此,但也可以有其它的解释,因为树种的分布除受到温度的影响外,还会受到其它环境因素如土壤、风速、湿度、pH、水流等的影响,植被的这种垂直分布格局不一定完全由温度解释。但是,毫无疑问,植物对温度的耐受性依然是塑造这种植被垂直格局的最重要的因素之一,特别是在温度类似于冻原的高海拔山顶。
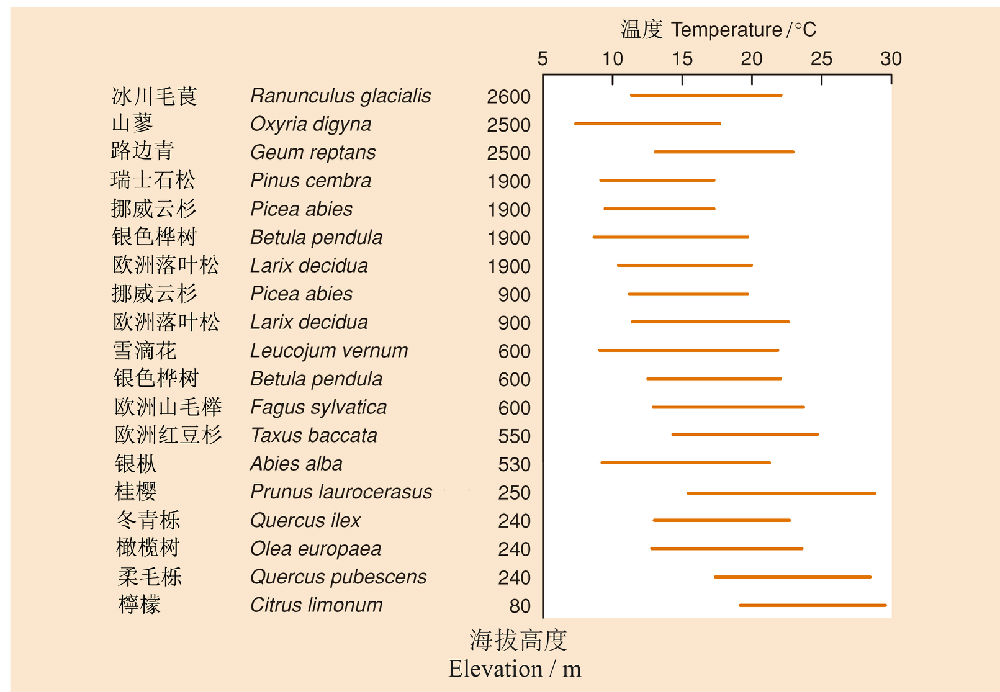
图9 欧洲阿尔卑斯山上能在低密度辐射(70Wm−2)条件下获得净光合作用的各种树种的温度范围(Begon et al. 2006引自Pisek et al. 1973)
三、热带-亚热带地区森林植被的地理格局与影响因素
在地球气候系统中的一个很大的特点之一就是在热带—亚热带气候区,在相对较窄的温度范围内,降雨呈现出巨大的变化,与中—高纬度地区相比,植被类型也最为多样(图3)。这里专门介绍降雨对(亚)热带地区植被类型的影响。
1. 南北半球降雨与干旱的时空格局
无季节差异的降雨仅发生在很窄的范围(赤道南北1o范围内)(图10)。南北半球热带-亚热带区域干湿变化的季节性格局也有相当程度的差异,总的趋势是,离赤道越远,气候越来越干燥,旱季时间越来越长,而且南北半球旱季(雨季)发生的时期则正好相反,譬如,南纬15o附近,旱季主要发生在5-9月,而在北纬15o附近,12月到次年的3月。
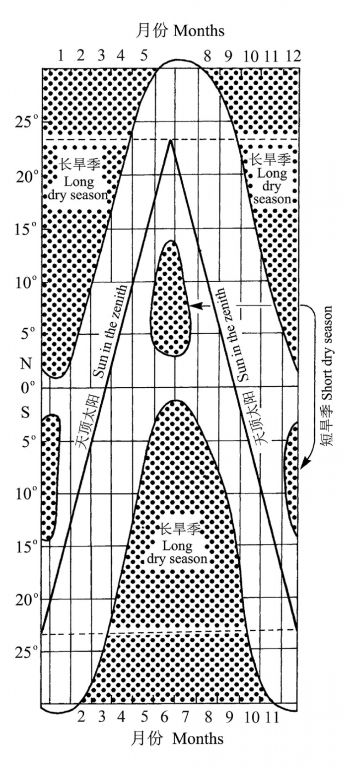
图10 热带亚热带的旱雨季及天顶降雨,阴影区域为雨季,点填充的区域为旱季(Lüttge 2008 引自Walter andBreckle 1984)
2. R-E指数(降雨量—蒸发量)显著影响植被类型
在热带-亚热带地区(类似的高温条件),降雨特征对森林植被的类型起着决定性的作用。一般用所谓降雨量与蒸发量之差(R-E指数)来表征气候的干湿,正值表明湿润的气候,而负值表示干燥的气候。图11显示了横穿委内瑞拉的7个站点的R-E指数与植被类型的关系,不难看出,在纬度10o(2→12o)的范围内,R-E值从接近+2000 mm变化降低到接近-3000mm,植被类型的变化变化巨大:热带雨林→常绿季雨林+湿润稀树草原→落叶林+西树干草原→干性落叶林→多刺灌丛。
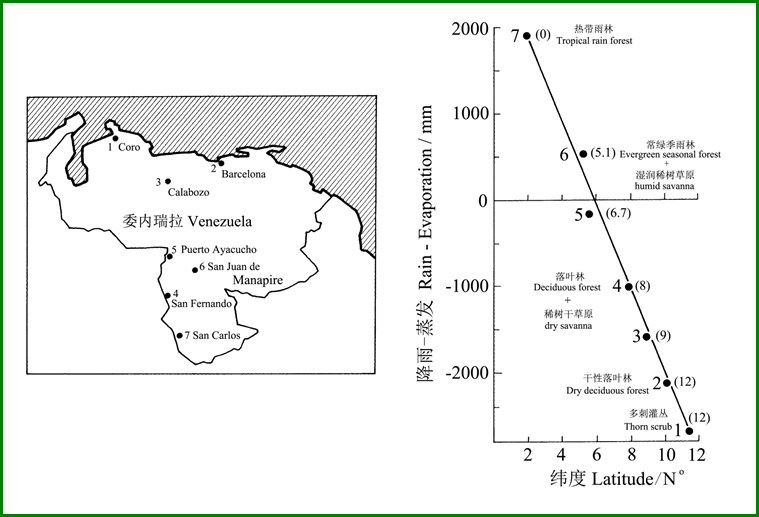
图11 横穿委内瑞拉的不同站点的降雨减蒸发量(R-E指数)与森林类型,在R-E指数后的括号中的数据表示每年干旱的月份(Lüttge 2008引自Medina1983)
3. 年降雨量和干旱期对森林类型的影响
以印度和委内瑞拉为例,依据降雨和干旱期的长短可以很好地区分不同的植被类型(图12)。降雨量与干旱期呈相反关系,降雨量大、干旱期短的区域生长雨林和季雨林,而降雨量小、干旱期长的区域则为刺灌丛、仙人掌林或沙漠。
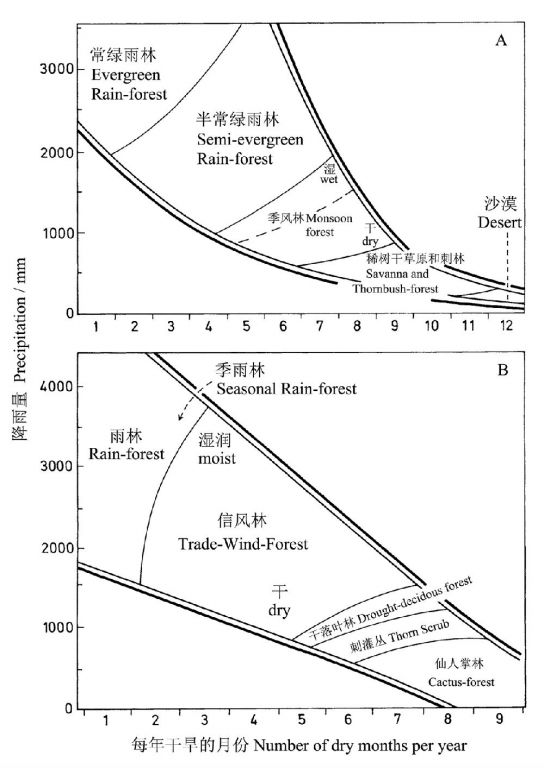
图12 与年降雨量和干旱期相关的印度(A)和委内瑞拉(B)的森林类型(Lüttge 2008仿Walter andBreckle 1984 and Vareschi 1980)
4. 降雨量对植物生活型的综合影响
沿热带梯度(降雨变化而温度相对稳定)植物生活型优势的变化如图5-13所示,主要变化模式为:(1)优势植被在最湿区域为高大的常绿树,而到了季节性干旱地区,则变为常绿和落叶混合林;(2)随着气候的进一步干燥,由于对关照的竞争小而对水分的竞争大,树和灌丛的高度下降,最终导致在干旱地区以多年生草本为主而灌木稀少的沙漠;(3)在极端干旱条件下,优势生活型变为一年生和球茎(在干旱季节,多年生草本地上部分死亡)。
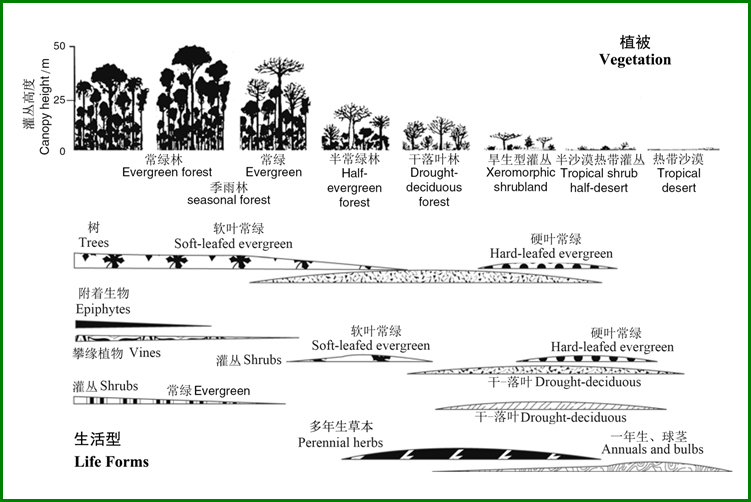
图13 沿热带梯度(降雨变化而温度相对稳定)植物生活型优势的变化(Chapin III 2011根据Ellenberg1979重新绘制)
四、气候—土壤—植被类型的耦合作用
土壤是所有陆地植物生存的基质,植物的养分也几乎来自土壤。虽然土壤的物理化学成分在一定程度上依赖于母岩的类型,但从大时空尺度来看,土壤类型(包括养分水平)是气候—生物群落生命活动联合塑造的结果,同时土壤环境又反过来作用与其上的生物群落。因此,植被类型也打上了土壤类型的印记。
1. 降雨—养分耦合作用对热带植被类型的影响
除了降雨外,养分对植被类型也有显著影响。图14显示了水分和养分对植被类型的协同作用,森林需要高的养分供给,或在养分较少的情况下,至少有充沛的水分,像生产力低的硬叶热带雨林。相反,在上述两类资源处于中—地水平的区域则被稀树草原所占领(Lüttge 2008)。

图14 依据养分和水分可利用性对各种稀树草原和热带森林的划分(Lüttge 2008引自Medina1987)
2. 气候对植被和土壤地带性格局的塑造
气候对植被的影响还与土壤发育密切耦合(图15):在靠近北极的极地荒漠和永久性冻土区,湿度和温度均很低,因此风化速率缓慢,有机质生产、分解和淋溶速率均十分缓慢,主要发育始成土;随着纬度的降低,湿度增高促进淋溶,而相对较低的温度导致相对较低的有机质分解,主要发育灰土、淋溶土和软土;随着纬度的进一步降低,虽然高温有助于化学风化,但由于蒸发大于降雨,低湿度阻碍植被的发育,土壤有机质积累甚少,主要发育干燥土;在高温和高湿的热带气候条件导致土壤快速风化和植物的生产力高,淋溶速率也快,而在较干旱的热带气候下,则发育稀树干草原,主要发育老成土、氧化土。
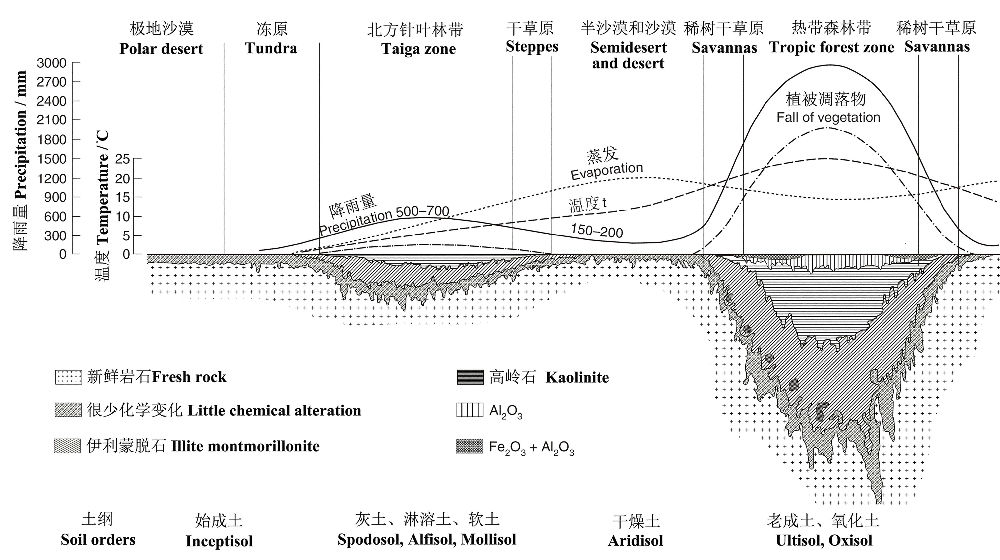
图15 沿着从赤道到北极的横断面,环境因子对陆地植物群系和土壤形成的影响(Paul 2007引自Birkeland1999)
3. 非地带性植被
一般来说,大气候决定土壤和植被类型的宏观地理格局,但是,即便是同样的气候类型,也不可能产生完全均一或单一的土壤类型。譬如,由于气候或地质或其它因素的影响,可能出现局域土壤类型的显著分化,出现一些十分特殊的土壤类型。
一种较为极端的例子是,同样的植被类型可以出现在几个不同地带中相似的土壤上,这种植被称为非地带性植被(azonal vegetation),如在一些特殊的土壤类型(如石质土、沙土、盐渍土、酸沼土、淹水的土壤、养分贫瘠或缺乏的土壤等)上,往往形成一些特殊的非地带性植被类型(有些类型分布还相当广泛),这些植被在很大程度上受土壤而不是气候的影响(沃尔特1984)。
生物量和生产力是表征植物群落生态功能的二个重要参数,物种相对多度则是表征植物群落结构的关键参数。在这里不探讨生态结构—功能参数的变化过程,只是比较其在地带性植被类型之间的差异。
1. 不同植物群系的生态功能
很显然,气候对植物群系的生产力和生物量有着深刻的影响,一般来说,温暖湿润的热带区域要远高于干旱或极端炎热或寒冷的区域(表5)。
表5-5 不同植物群系的净生产率和生物量
|
Types of biomes |
NPP范围 NPP range (g/m2) |
平均NPP Average NPP (g/m2) |
生物量Biomass (kg/m2) |
平均生物量 Average Biomass (kg/m2) |
|
热带雨林Tropical forest |
1000-3500 |
2200 |
6-80 |
45 |
|
温带落叶林Temperate deciduous forest |
600-2500 |
1200 |
6-60 |
30 |
|
针叶林Taiga |
400-2000 |
800 |
6-40 |
20 |
|
灌木Chaparral |
250-1200 |
700 |
2-20 |
6 |
|
稀树大草原Savannah |
200-2000 |
900 |
0.2-15 |
4 |
|
大草原Prairie |
200-1500 |
600 |
0.2-5 |
1.6 |
|
冻原Tundra |
10-400 |
140 |
0.1-3 |
0.6 |
|
沙漠Desert |
10-250 |
90 |
0.1-4 |
0.7 |
(数据源自Odum and Barrett 2005)
2. 不同植物群系的物种相对多度的比较
Hubbell(2001)比较了4种不同森林类型(群系),即北方林(一种温带亚高山林,位于美国大雾山国家公园克凌曼圆顶的山顶,样方面积=0.2 ha,少于10个物种)、温带落叶林(位于大雾山国家公园的低海拔高度地区,样方面积=1 ha,大约40个物种)、热带半落叶林(位于哥斯达黎加,样方面积=13 ha,约120个物种)和热带长绿林(位于巴西,样方面积=4 ha,大于200个物种)的种相对多度曲线。这四种森林生态系统的物种多样性和生产力均为:北方林<温带落叶林<热带半落叶林<热带常绿林。
从种相对多度曲线(图5-16)可以看出,当森林生态系统从北方林→温带落叶林→热带半落叶林→热带常绿林变化时,种相对多度的模式由近乎于直线转变为S型曲线,即第一优势树种的优势地位下降,而中等优势度的树种显著增多。这与后面要介绍的温带地区弃耕地向森林生态系统的演替过程有许多类似之处。
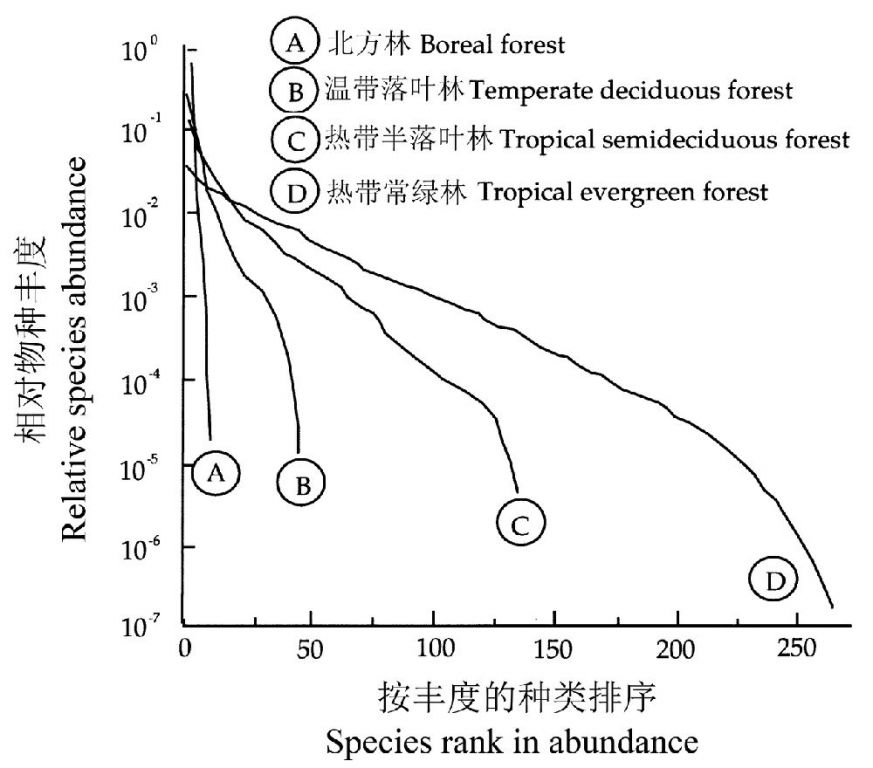
图16 跨越大的纬度梯度的四种郁闭林的树种多样性—优势度曲线(引自Hubbell 1979, 2001)
六、历史因素对植被类型的影响
气候虽然是决定植被类型的重要因素,但不是唯一因素。由于生命进化的随机性,加上地质历史原因(譬如板块运动引起的隔离历史的不同),即使同样的气候背景,不同地理区域的植物区系也许向完全不同的方向进化,因此可能演化出完全不同的植被类型。
一个典型的例子就是肉质植物,在美国的干旱地为仙人掌科(Cactaceae),但在非洲却为大戟科的大戟属(Euphorbia),这在植物生态学称之为趋同现象,而在气候相似的澳大利亚,无论在哪里都见不到肉质茎植物;但是,澳大利亚的植被在群落外貌上也十分不同于其它大陆,甚至其哺乳动物区系也是独一无二的,这可能与澳大利较早脱离亚冈瓦那古陆有关。此外,在泛北极植物区普遍分布的落叶林,在新西兰温带气候条件下却完全不存在;又如,在系统发育上比较古老的针叶树类群,罗汉松科和南洋杉属仅限于南半球分布,而种类繁多的松科和几乎所有的杉科植物仅限于北半球分布,但是柏科却遍布所有大陆(沃尔特1984)。
物种的地理分布格局并不一定都与其绝对分布极限因子相重合,换句话说,一个物种的自然分布并不是反映其生理需求的绝对指南(沃尔特1984)。当然,也确实存在一些物种的分布可能达到其极端环境(温度、湿度、盐度、光照等)因子的限值范围。其实,可能大多数物种由于历史地理的或生态的原因远未达到其分布的自然界限,因此,如果人们有意或无意地将一个物种移入另一个未曾分布的区域,经常会带来外来物种缺乏天然的控制机制(如天敌)出现灾难性增长,这就是现在广为关注的外来种问题。
结语
绿色植物是地球上所有生命的源泉,植物学家创造了一系列的名称来描述生活在一定空间中的植物集群—植物群落、生活型、植被、生物群系、植物区系等,其中植被泛指地表所覆盖的所有植物,它基于外貌的分类体系也使非植物学家的人士容易理解。
从本质上来讲,地球上植被的现代地理格局不仅是现代气候(主要是温度和降雨)格局的产物,更是气候变动与生命系统在漫长的地质历史过程中相互作用的产物。无可争辩的事实表明,在地理尺度上,气候几乎彻底地主宰了植被的大尺度空间格局。这一点在进化生物学上十分重要,因为它在宏观上决定或约束了物种、群落乃至生态系统的演化方向与基本格局,虽然还必须立足于一定的生物地质历史背景的基础之上。植被地理格局的存在也暗示物种(因此群落)的进化与分布并不那么随机,它被气候的格局和历史宏观性地决定,虽然不可避免地伴随局域与暂时的随机性。也可以说,它指明了微观进化的随机性中蕴藏的宏观方向性。
植被地理格局的存在也昭示了气候环境约束下的植物群落的自我组织、自我调节、自我更新与自我发展—即方向性演替模式的存在,同时也昭示植物的进化与植被的演替必定是偶然(随机)性与必然性相互作用的产物,而且偶然与必然在一定程度上互相补充、互为因果,即必然性一方面约束系统的演化,而另一方面它自身又是演化的结果,它们不可分割。此外,这种空间格局还昭示了宏观的生命世界演化历史的存在,它使过去、现在和未来得以串接与融合。
参考:从生态学透视生命系统的设计、运作与演化—生态、遗传和进化通过生殖的融合(谢平著,科学出版社,2013)
电子版下载:http://wetland.ihb.cas.cn/lwycbw/qt/
联系方式:沈宏 徐军 027-86780056 Email:wetland@ihb.ac.cn
地址:武汉市武昌区东湖南路7号
访问数量:
