 当前位置:
当前位置:(本文来源:http://blog.sciencenet.cn/blog-1475614-821037.html)
“演替”是在生态学发展早期提出的用于解释植物群落时序—方向性变化与更替机制的核心概念之一,关乎到人类对植物资源的合理利用。在相同的气候模式(即中、短时间尺度)下,陆地上的植物群落(植被)如何演替呢?从总体上来看,植被演替伴随着生态系统中有机质的不断累积,物种多样化,关系复杂化与结构层次化。这似乎与英国洛桑实验站草地持续施肥的试验结果(见博文:英国人用百年生态试验换来一个植被演替的"谬论"?)背道而驰......
植物群落的变化规律一直都是植物生态学研究的主要问题之一。植物群落的变化幅度会有所不同,一般将具有一定方向性和时序的物种组成的变化—即一个(类)物种被另一个(类)物种所取代—过程称为演替(succession),而将较小程度的物种组成的变化(未出现物种更替)过程称为波动(fluctuation)。因此,演替一般发生在相对较长的时间尺度上,而波动发生在相对较短的时间尺度。演替也可看作是植物群落结构在时间尺度上的序列关系。
植被的演替可区分为两种基本类型:原生演替和次生演替。原生演替对了解植被的发展过程十分重要,但是在现实世界中却很少被观察到,因为它们进展缓慢,往往可跨越数世纪以上。人为或自然的扰动(如火烧、耕作等)可对原始植物群落产生破坏,在干扰消除后,植物群落发生所谓的“次生演替”过程,这通常是一种较短时间尺度(数十年)的生态过程。
一、原生演替—需要改造基质的缓慢过程
在一块原生裸地(暴露的岩石)上,经过一系列的变化在原来不曾有过植被的裸露基质上逐渐建立起植物群落(图1)。以花岗岩石上的原生演替为例,说明原生演替的大致过程。在乔治亚的山麓地带到处可见到露出地面的花岗岩石,原生演替起始于花岗岩表面由于侵蚀出现的凹陷处,能在裸露岩石上的干热条件下生存的地衣首先进入,随着沙土和有机质的积累,能在薄土上生长的一年生植物(如景天、驯鹿苔藓)定居开始定居,随着土壤层的变厚,真正的苔藓和一年生草本开始建群。随着土壤保水能力的增加,多年生草本开始出现,最后,植被和足够的土壤堆积支撑多年生木本植物(如厚皮刺果松、西部红松、白莓)的生长。在花岗岩石上的原生演替十分缓慢,据估计从裸露的花岗岩石转变到松树灌丛需要700年以上(Gibson& Gibson 2006)。

图1 在原生演替过程中,经过一系列的变化在原来不曾有过植被的裸露的基质(如暴露的岩石)上建立起植物群落,这一过程缓慢,需要很多年(引自Gibson & Gibson 2006)
二、次生演替—扰动后较快的群落重建过程
1.草原次生演替
热带稀树草原(委内瑞拉卡)火烧后的次生演替
在委内瑞拉卡拉沃索(Calabozo)进行的实验表明,在最后一次火烧后,被保护区域的稀疏草原的地上总生物量在最初的4年间不断增加,然后稳定在一定的水平,其中绿色生物量变化不大,而干生物量不断在最初的4年间不断增长(图2)。
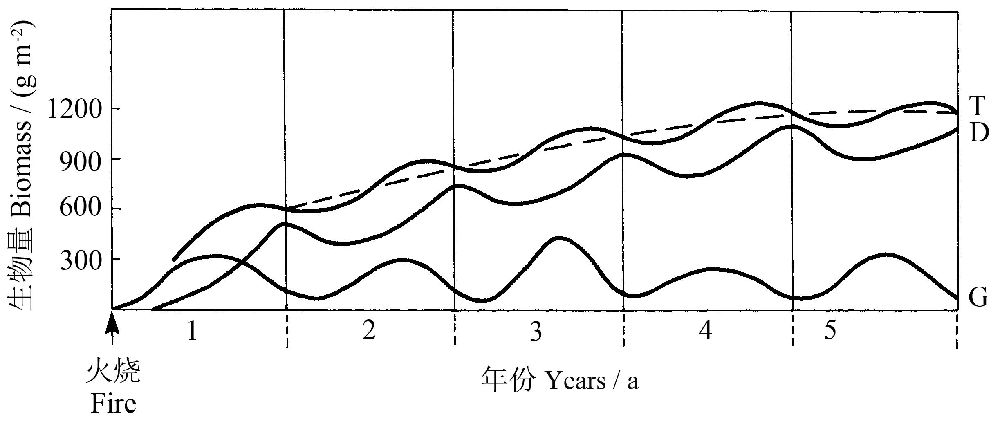
图2 委内瑞拉卡拉沃索一稀树草原地上生物量(T为总生物量,G为绿色生物量,D为干生物量)的变化,火烧经过5年后达到稳定状态(虚线)(Lüttge2008引自Sarmiento 1984)
温带沙原(美国)弃耕后的次生演替
人们将森林或草地开垦为农耕地,后又当耕地被废弃后所发生的次生演替,称为弃耕地演替(或撂荒地演替)。Inouye等(1987)调查了美国明尼苏达州锡达溪自然历史区域(该区域在西部的草原和东部的落叶林之间)位于沙原上的22块弃耕地(弃耕时间从1-56年不等)的植被,发现随着弃耕年龄的增加,土壤N含量、植被覆盖率、植物总地上生物量和枯落物盖度均显著增加。Tilman& Wedin(1991)根据他们的数据结合其它资料,绘制了次生演替过程中7个优势种(肯塔基蓝草Poa pratensis,匍匐冰草Agropyron repens,豚草Ambrosia artemisiifolia,剪股颖Agrostis scabra,帚状裂稃草Schizachyrium scoparium,大须芒草Andropogon gerardii,一种悬钩子Rubus sp.)的相对丰度的变化图,认为演替后期的种类对N的竞争力更强(图3)。与热带稀树草原相比,演替达到平衡的时间要长得多—至少半个世纪或以上。
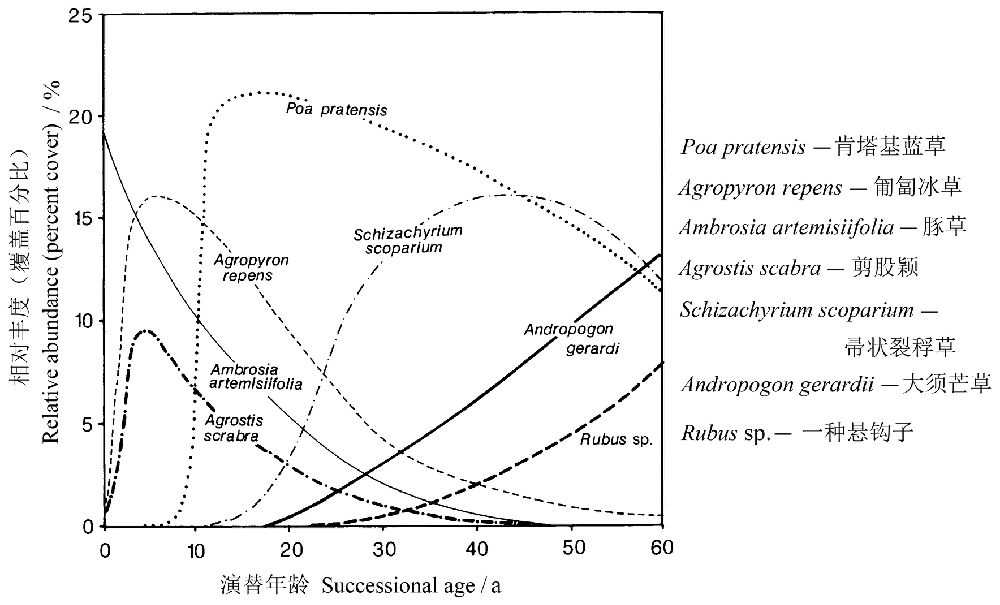
图3 美国锡达溪自然历史区域的演替,次生演替过程中7个优势种的相对丰度是根据22块弃耕地的年代序列数据(Inouyeet al. 1987)和其它观测数据计算得来的(引自Tilman& Wedin 1991)
2.森林次生演替
火烧是干燥的森林和草地中常见的一种自然现象,据称它的一个重要的生态功能就是可以替代分解者将积累的枯枝落叶迅速地矿质化。由闪电引起的火灾甚至在石炭纪的森林中就曾发生过!自然火烧已是许多生态系统(如干旱期的草地、冬雨区的疏林地、针叶林地等)的一种正常甚至是必需的生态过程(沃尔特 1984)。
火烧引起的森林演替(称之为林分替换火)受到广泛关注,林分火能显著影响种类的组成。为了使某一物种适应一定火烧周期生存下来,在该物种充分成熟到能繁殖之前(不然没有种源)火烧不能发生,但是在大部分老龄个体死亡之前(被后面演替的种类取代之前)必须发生。针对火烧的轮换周期(Rotation period)指经过林分火后植被的平均恢复间隔(Frelich 2002)。
在北美的大湖区(温带大陆性气候),树种可划分为两类:1)火烧依赖型种类—在火烧周期在300年或更短时变为优势;2)火烧非依赖型(常常对火烧敏感)种类—火烧周期长于500年时变为优势(图4)。有一个种类,黑云杉,能适应上述二种情形,因其在大湖区的树种中具有特殊的能力无论在短的还是在长的火烧周期中都能很好地繁殖。
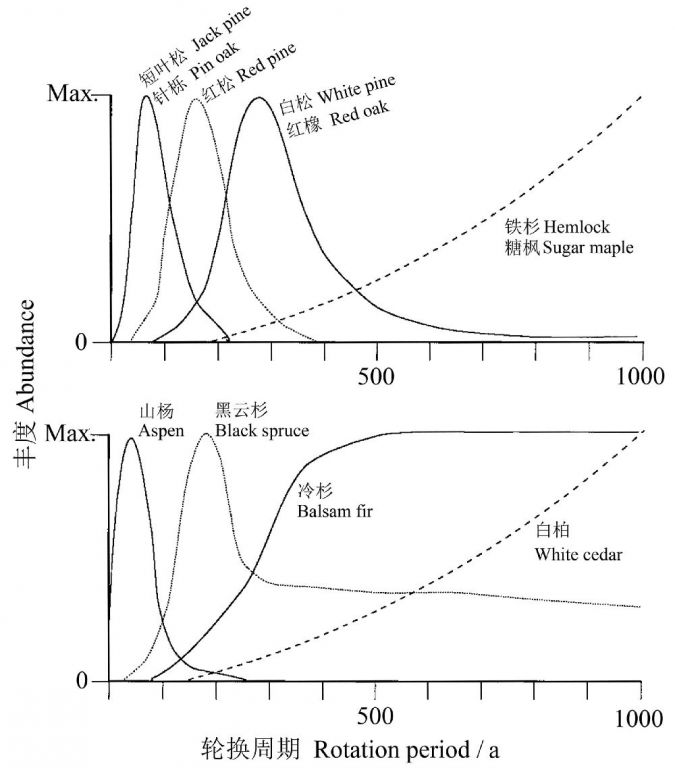
图4 大湖区林分替换火的轮换周期与一些重要树种丰度的关系。y轴表示相对丰度(刻度无单位),假设每个种在某个轮换期达到最大丰度(引自Frelich 2002)
三、演替—伴随着一系列生态系统特征的变化
Odum(1969)以森林生态系统为例,从群落能量学、群落结构、生活史、养分循环、选择压力、内稳态等方面勾勒出生态演替过程中生态系统特征的变化趋势(表1)。虽然只是定性的描述,但是得到广泛认同。
Odum(1969)图解了森林生态系统演替过程中主要能量学参数的变化趋势(图5),PG和R之差为净初级生产量,在演替早期(或青年期),群落的总初级生产量超过呼吸量,因此P/R大于1(在有机污染的特殊情况下,可能出现P/R<1)。临近群落演替的后期(或老龄期),P/R→1,即在“顶级”的生态系统,能量的固定和消耗趋于平衡,因此,P/R系数是表征系统相对成熟度的很好的功能性指数。
表1 生态演替过程中预期的生态系统变化趋势
|
生态系统特征 |
发展期 |
成熟期 |
|
群落能量学Community energetics |
|
|
|
1. 毛生产量/群落呼吸(P/R) |
大于或小于1 |
接近1 |
|
2. 毛生产量/生物量(P/B) |
高 |
低 |
|
3. 单位能量流动支撑的生物量(B/E) |
低 |
高 |
|
4. 净群落生产量 |
高 |
低 |
|
5. 食物链 |
线形,牧食食物链占优势 |
网状,碎屑食物链占优势 |
|
群落结构 Community structure |
|
|
|
6. 有机物总量 |
小 |
大 |
|
7. 无机营养盐 |
生物外 |
生物内 |
|
8. 物种多样性—组分变异 |
低 |
高 |
|
8. 物种多样性—均匀度 |
低 |
高 |
|
10. 生化多样性 |
低 |
高 |
|
11. 分层和空间异质性(格局多样性) |
缺乏组织性 |
组织程度高 |
|
生活史 Life History |
|
|
|
12. 生态位特化 |
宽 |
窄 |
|
13. 有机体体积 |
小 |
大 |
|
14. 生命周期 |
短、简单 |
长、复杂 |
|
养分循环 Nutrient cycles |
|
|
|
15. 矿物质循环 |
开放 |
封闭 |
|
16. 养分交换速率(在有机体和环境之间) |
快 |
慢 |
|
17. 碎屑在养分再生中的作用 |
不重要 |
重要 |
|
选择压力 Selection pressure |
|
|
|
18.生长型 |
快速生长(r-对策) |
反馈控制(K-对策) |
|
19. 生产 |
量 |
质 |
|
内稳态Overall Homeostasis |
|
|
|
20. 内共生 |
不发达 |
发达 |
|
21. 养分保存能力 |
弱 |
强 |
|
22. 稳定性(对干扰的抗性) |
弱 |
强 |
|
23. 熵 |
高 |
低 |
|
24. 信息 |
小 |
大 |
(引自Odum 1969)
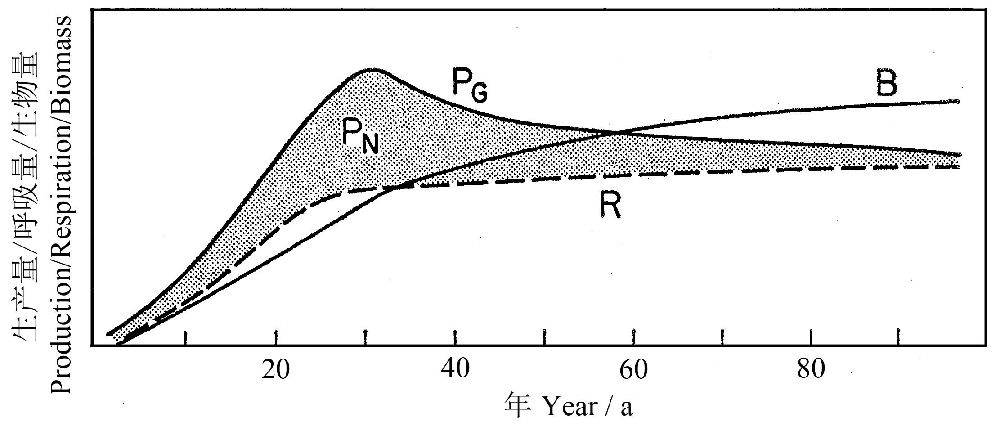
图5 一个森林演替过程中能量学参数的比较。PG, gross production; PN—净产量, R—群落呼吸量,B—总生物量(引自Odum1969)
Dierschke(1994)以在亚大西洋气候条件下硅酸盐岩石上起始的原生演替为例,分析了演替过程中生态系统主要特征的动态变化趋势,在演替的起始阶段,不可预测的杂乱的相互作用占优势,因为在很大程度上,繁殖体的侵入和建群是随机的,广布性一年生植物占优势。随后,优势植物群落依次经过多年生禾草→灌丛→森林的循序演替(图6)。
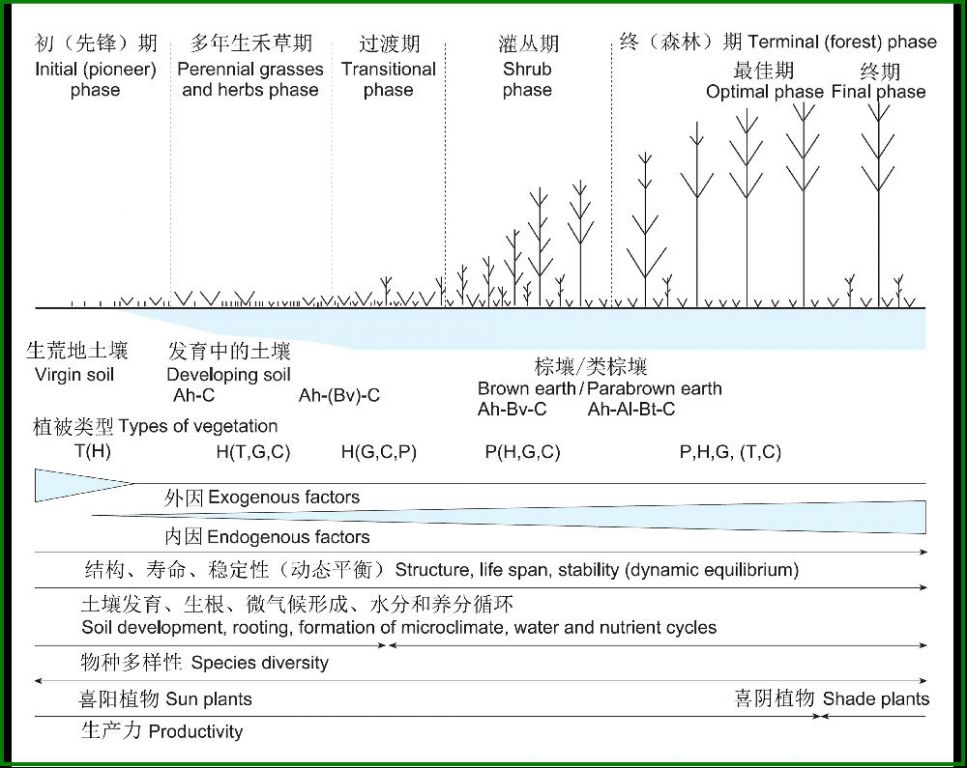
图6 原生演替过程中的主要特征(在硅酸盐岩石以及亚大西洋气候条件下)(Dierschke 1994,间接引自Schulze 2005)
四、次生演替的过程—接力植物区系假说和初始植物相组成假说
Egler(1954)提出了演替的二种可能情景,一种称之为接力植物区系假说(relay floristics),也主要是Clementsian的观点,即协调的群落像接力赛跑一样一个取代另一个(图7A);另一种他认为更可能,称之为初始植物相组成假说(initial floristic composition hypothesis),强调定居的过程以及物种生命周期的差别(图7B)。Egler以美国东部弃耕地的演替为例,认为所有种类在演替的早期就到达了,早期的种类像一年生和多年生草本因为生长快首先占据优势,树种虽然也早就到达,但直到很后才占据优势,是因为生长慢。因此,他认为特定的演替系列是初始植物群落组成差异的直接结果。
上述二个假设的最大区别就是“是否所有种类在演替初期就到达”,其实在偌大的一个地球系统,涵盖几乎无数的生境类型及极端多样的环境条件,各种次生演替的类型、程度和规模也都千差万别,上述二种类型以及二者不同程度的混合情况都有可能存在。
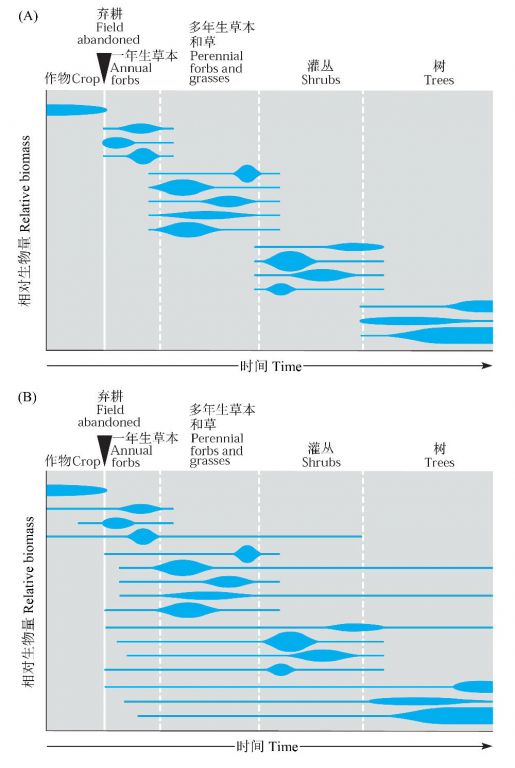
图7 在一个假想的北卡罗莱娜弃耕地上仿Egler(1954)的理论绘制的演替简图。每条线代表所属植被类型的一个种类,线越宽,表示在给定时间该种越重要。(A)根据接力植物区系假说,像接力赛上的选手一样一个类群取代另一个,(B)初始植物相组成假说,为Egler的接力植物区系假说的修改,这里,所有的种类在演替的初期就出现,演替仅是各种生活史的一个伸展过程(引自Gurevitch et al. 2002)
五、带森林区弃耕地的演替—物种相对多度从直线转变为S型
物种相对多度的变化也是刻画生物群落的重要指标之一,它可以用来判断植物群落的演替阶段。图8为美国伊利诺斯南部(温带气候)五个不同弃耕阶段的撩荒地中,植物群落物种相对多度的变化。在演替的早期阶段,植物群落常常由少数最先到达的杂草(先锋种)所占据,这些先锋种往往是一些能快速生长的r—对策种,然后逐渐被灌木和乔木群落重新占领。有意思的是,随着演替的进展,种相对多度的模式由近乎于直线转变为S型曲线,中等优势度的物种显著增多。

图8 在伊利诺斯南部,五个不同弃耕阶段的撩荒地中,种相对多度的模式。模式表示为在一个群落中一给定种占全部种所覆盖的总面积的百分数相对于种的等级(按从多度最大到最小的顺序)绘出曲线。空符号代表草本植物,半空的代表灌木,实的代表乔木(引自Bazzaz 1975)
Lee等(2002)研究了位于韩国中部的京畿道(温带气候)地区的水稻弃耕地的次生演替,所选的研究地点分别代表了5个不同的演替阶段:刚弃耕、弃耕后3、7、10年以及50年的日本桤木森林。随着弃耕年份的增长,植被优势种出现明显的更替:看麦娘Alopecurusaequalis(一年生杂草)→竹头草Aneilemakeisak(一年生草本)→灯心草Juncuseffuses→尖叶紫柳Salixkoriyanagi→日本桤木Alnusjaponic,最终植被形成稳定的桤木森林群落,而桤木群落被认为是与区域性气候背景下的顶级群落(图9)。随着演替的发展,物种多样性逐渐增加,土壤有机质、N、P、K、Ca、Mg也不断增加。
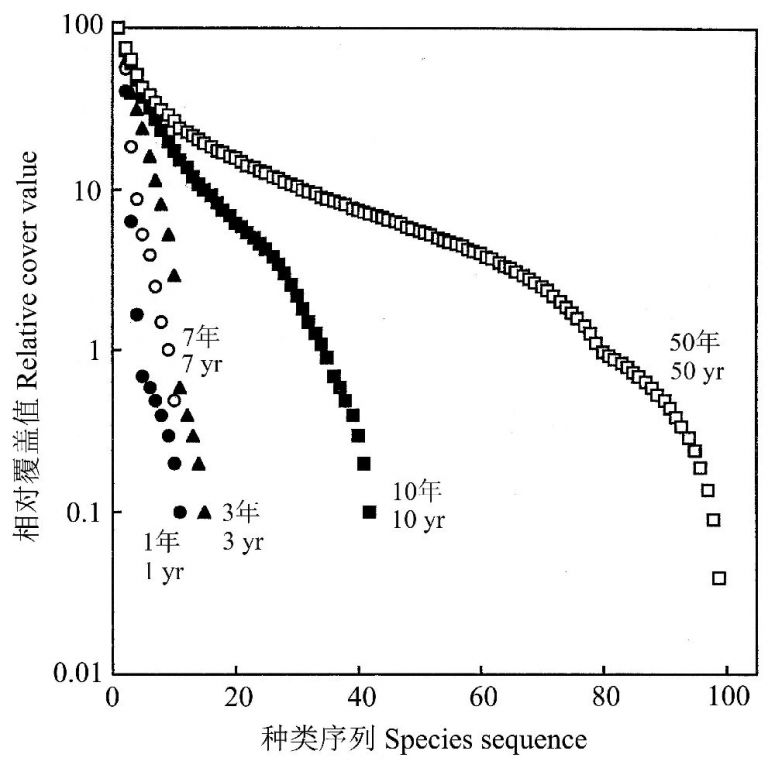
图9 各种年龄弃耕地的植物物种排序-丰度曲线,重要性值根据5个弃耕年龄组的56个样方的相对覆盖估算值求得(引自Lee et al. 2002)
结语
原生演替花费漫长的时间在适合于后期植被发展的土壤及其有机质的形成与积累的基础之上,发展到一定程度后,与次生演替过程相似。这种中短时间尺度上的演替也能看成是一种时序性生态同化,它是一种适应性的群落结构发育、更替与融合过程,从总体来看,植物群落趋于(物种)多样化、(关系)复杂化、(结构)层次化、以及(状态)稳定化。它也是一种环境(主要是气候)约束型的自组织式的结构发育过程。
这种时间尺度的演替基本上不涉及物种分化问题(成种事件需要更长的时间),因此,演替(无论是原生还是次生)过程只是一种物种的迁入或再迁入以及随之而来的物种间的相互作用与相对平衡的形成过程。植被演替本质上是地质历史变动轨迹中植物群落的一种气候约束性或归宿性反应,它与地域性植被类型以及影响扩散的因素(如地貌)密切相关。
参考:从生态学透视生命系统的设计、运作与演化—生态、遗传和进化通过生殖的融合(谢平著,科学出版社,2013)
电子版下载:http://wetland.ihb.cas.cn/lwycbw/qt/
联系方式:沈宏 徐军 027-86780056 Email:wetland@ihb.ac.cn
地址:武汉市武昌区东湖南路7号
访问数量:
