 当前位置:
当前位置:(本文来源:http://blog.sciencenet.cn/blog-1475614-822102.html)
云谲波诡、风花雪月、草木轮回、沧海桑田……尘世间万物皆变,植被亦如此。演替—植物群落结构在时间尺度上的序列关系,演替—地质历史变动轨迹中植物的归宿性反应……它有方向,或有终点?是什么驱动森林或草原像云雾式地消散或折回?
“演替”这个植物学上的核心概念自18世纪开始就受到人们的关注,演替模式(轨迹)·机制也是人们长期探索的问题,倾注了无数科学家的心血,因此也诞生出了各式各样的概念、模型或理论,但也不乏争议。
一、在时空尺度中丈量演替
离开空间尺度来谈论植被的时间变化规律几乎是没有意义的,因为植被的变化必须耦合在时空尺度之中!一些植物地理学或植物生态学家早已关注这样的问题。
图1显示植被的变化跨越巨大的时空尺度,为了方便起见,可将从短到长的变化依次称为波动(Fluctuation)、林隙和斑块动态(Gap and patch dynamics)、周期性演替(Cyclic succession)、次生演替(Secondary succession)、原生演替(Primarysuccession)和久期演替(Secular succession);而在空间上,从个体→种群→群落→景观→群系→整个生物圈!
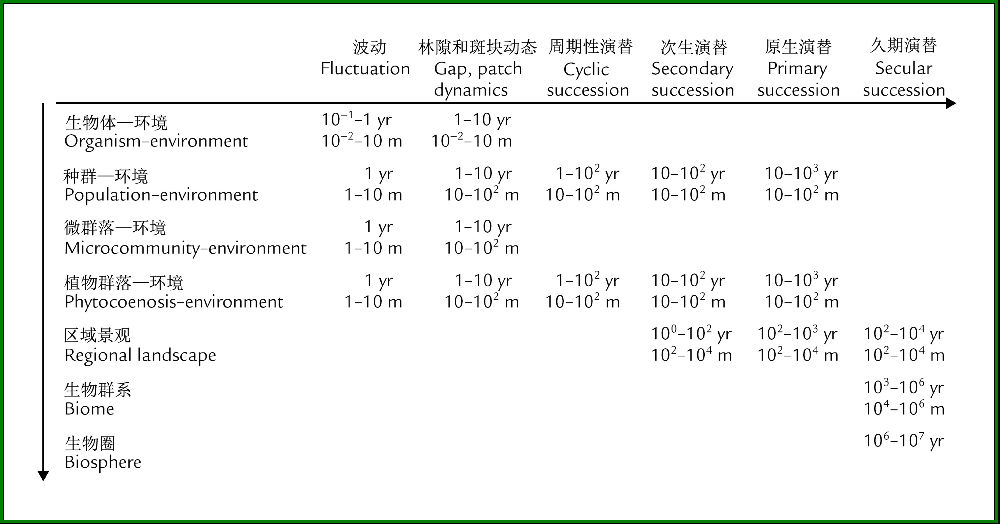
图1 植被动态变化的空间(m)和时间(yr)尺度(引自van der Maarel 1996)
二、演替理论之来龙去脉
从某种意义上来说,长期的生态演替(Secular ecological succession)早在18世纪就曾引起林奈(Linnaeus)、布丰(Buffon)以及在19世纪引起奥古斯丁·彼拉姆斯·德·堪多(Augustin-Pyramus de Candolle)、洪堡德(Humboldt)、莱尔(Lyell)等的注意(Lawley 2009)。19世纪初,法国博物学家Dureau de la Malle(1825)首先使用了演替一词来描述森林砍伐后植被的发展过程。
1860年,美国哲学家和自然主义者亨利•梭罗(Henry David Thoreau)在其撰写的“The Succession of Forest Trees”一文中,从直觉的科学观点阐述了生态演替是如何进行的,认为动物和气象(风、雨水)对树木种子的搬运作用扮演了重要角色,较轻的种子(如松树和枫树的种子)主要通过风和雨水搬运,而较重的种子(如橡子)通过动物搬运。譬如,通过松鼠搬运橡子,使松树被砍伐后橡树得以取而代之,而后地下的松子又可使松树取代橡树。
十九世纪末,美国植物学家Cowles(1899)研究了Michigan湖岸的沙丘上的植被的发展过程,进一步发展了演替的概念。他阐述了原生演替及其序列(在特定环境下一系列可重复的群落变化)的思想,正如他所指出的“The ecologist, then, must study the orderof succession of the plant societies in the development of a region, and hemust endeavor to discover the laws which govern the panoramic changes”。
二十世纪初,美国植物学家Clements(1916)首次将演替描述成由下述六个基本过程所组成:
1)裸化(Nudation):演替起始于裸地(如由于扰动)的发展;
2)迁移(Migration):植物繁殖体的到达;
3)定居(Ecesis):建群和植被的初期生长;
4)竞争(Competition):当植被开始建立、生长和扩张时,各物种开始为空间、光照和营养竞争,导致一种植物被另一个所取代;
5)相互作用(Reaction):植物的生长和死亡影响栖息地环境,而这反过来又影响资源的可得性;
6)稳定(Stabilization):这种相互作用最终将导致顶级群落的出现。
这一描述性的演替理论对后来的生态学思想产生了巨大的影响,被认为是一个经典的生态学理论,Clements也被称为植物群落演替理论的奠基人。
不久,美国植物学家Gleason(1927)对演替的概念重新进行了讨论,提出了与Clements明显不同的观点,认为演替更复杂,更具不确定性,随机因素的作用更大,否认具有一致的、清晰分界的群落类型的存在。
三、五花八门地划分演替
人们为了从各种角度认识演替,依据不同的原则对演替进行了分类。常见的,可按基质性质、生态系统层次、驱动因素、经历的时间以及生态系统类型等来划分演替的类型如下:
1.按演替起始时的基质性质可分为以下二类:
原生演替(Primary successions)—指发生在原生裸地(barren land)上的演替。所谓原生裸地就是从来没有植物覆盖的地面(如新暴露的岩石、沙丘等),或者是原来存在过植被,但被彻底消灭了(包括原有植被下的土壤)的地段。
原生演替一般起始于一些新形成的(如火上喷发)裸地,也可以说是从没有生命体的一片空地上开始的植被类群的演替。现在地球上所有的动植物和土壤都是原生演替的产物(Walker & Moral 2003)。图2是发生在沙丘上的原生演替。

图2 在沙丘上的原生演替。滨草是最先建群的植物,它可稳定沙丘,使灌木和树能生长(引自Miller & Harley 2001)
次生演替(Secondary succession)—指发生在次生裸地(secondary barren)上的演替。所谓次生裸地是指原有植物群落被破坏(如森林砍伐、火灾、洪水等),但原有植被影响下的土壤条件仍然存在或受到很少破坏,甚至还残留原有植被的种子或繁殖体的裸地。也有一些学者认为次生演替也可称之为再生演替(regeneration succession),因为确实是生物群落的再生(如van der Maarel 1988)。图3是森林砍伐形成的次生裸地。

图3 砍伐前(A)和砍伐后用于耕作(B)的巴西热带雨林。土壤很快贫瘠,然后弃耕,再转移到邻近的森林。弃耕后便开始次生演替(引自Miller & Harley 2001)
2.按生态系统层次可分为:
种类演替(Species succession):这一词语在(无论陆生还是水生)生态学论文中用法十分广泛,但仅指种类的更替而已。
种群演替(Population succession):也有一些学者使用这一词语,如“bird population succession”、“microbial population succession”等,含有种群数量信息(如优势种群)的种类演替的用法;此外,也被用于社会科学中与人口相关的描述,如“human population succession”。
群落演替(Community succession):是指植物群落发展变化,一个群落代替另一个群落的自然演变现象;演替具有方向性,即由低级到高级,由简单到复杂,一个阶段接着一个阶段。
生态系统演替(Ecosystem succession):也有一些学者使用“生态系统演替”的概念,譬如Gutierrez 和Fey(1980)在其专著“生态系统演替”中这样描述道:“自然生态系统如湖泊、草地和森林从其发展早期向成熟演替,此时动植物群落与其物理环境达到一种平衡状态”。其实,生态系统演替在本质上就是我们前面讲述的生态系统稳态转化。
一般来说,用生态演替(ecologicalsuccession)泛指以群落演替为主的演替的用法最为广泛,当然,群落演替的用法也十分广泛,而生态系统演替的用法并不常用。而种类演替、种群演替基本不是通常意义的演替。
3.按演替的驱动因素可分为:
群落发生演替:指在原生或次生裸地上植物群落入侵与发生的过程;
内因生态发生演替(或内因动态演替):由植被自身变化驱动的演替。譬如,植物的生长可导致土壤有机质的积累,改变土壤的养分和pH,从而驱动群落的演替。
外因生态演替(或外因动态演替):由外部环境影响(而非植被本身,如动物活动与牧食、火山喷发、洪水、火烧等)驱动的演替。
地因生态演替:由于大范围内统一的变化所导致植物群落变化的过程,如整个区域地理环境的改变所引起的演替。
这种按演替驱动因子来划分群落演替类型是前苏联植物学家Cykaqëb BH于二十世纪中叶提出的(曲仲湘等1984)。
其实,群落发生演替可以理解为演替的早期,此阶段植物对环境的改变作用有限,而内因生态演替可以理解为演替的后期,植物的生命活动显著改变土壤环境,而外因生态演替则是较强烈的环境(或周期性)扰动的结果,应该还未严重到形成原生或次生裸地的程度。而地因演替则是指大时空尺度(地理)环境变化驱动的植被变化。
4.按演替所经历的时间长短可分为:
快速演替:在数年乃至十几年间所发生的演替;
长期演替:数十年甚至达百年的演替;
世纪演替:在地质年代尺度上的演替。
5.按生态系统类型可分为:
水生演替系列(Hydrosere)
石生演替序列(Lithosere)
沙生演替序列(Psammosere)
旱生演替序列(Xerosere)
盐生演替序列(Halosere)
四、困惑—演替的轨迹与终点
自生态科学存在以来,森林群落演替的方向性和终点就是一个关注的主要焦点之一。其中,以顶级群落理论为代表的学说认为植物群落的演替最终将会趋于稳定群落。
1.演替终点
气候顶级学说:Clements(1916)最早使用顶级(climax)这一词语,提出了单一的气候顶级(Climatix climax)的思想。他认为,某一区域内的植被演替的终点取决于该地区的气候条件,将会出现与气候条件相适应的优势群落,在气候条件相对稳定、没有其它干扰的情况下,不会出现新的优势植物,即形成所谓气候顶级群落,也称为单顶级(Mono-climax)。根据这一学说,一个气候区只有一个潜在的气候顶级群落,只要经过充分的时间,该区域的任一生境最终都能发展到这种群落。尽管Clements的气候顶级学说备受争议,但这一学说对后来的生态学思想的强烈影响仍然长达半个世纪之久(Selleck 1960)。
多顶级学说:在Clememts气候顶级学说的基础上,Tansley(1939)提出了多顶级(polyclimax)学说,认为一个气候区域内可以有多个顶级群落,除了形成气候顶级之外,还可以有土壤顶级(edaphic climax)、地形顶级(physiographic climax)、由放牧或动物啃食形成的生物顶级(biotic climax)、由人类活动维持的人为顶级(anthropeic climax)以及演替偏向了一个新的线路因此维持在一个新的平衡下的偏途顶级(plagioclimax)等。也就是说,气候顶级是气候在较大范围内控制植被演替的结果,而其它演替类型则来源于除气候外的其它因素在较局域的尺度上对植被演替的控制。
顶级群落格局学说:Whitaker(1953)通过对沿环境梯度的种群分布的分析,提出了顶级群落格局假说(climax pattern hypothesis),他提出用与环境梯度相应的逐渐变化的群落格局来解释多顶级现象,认为格局中心分布最广的群落类型就是气候顶级。他建议用可定义的优势顶级(prevailing climax)还取代不可定义的气候顶级。所谓优势顶级群落指占据最大区域的植被类型。
这三个学说既有联系又有区别,也是人们对演替终点认识不断深化的过程。气候顶级应该真实地反映了大尺度的气候条件对植被演替终点的影响,而多顶级学说补充了局域尺度的植被演替终点的控制机制。顶级群落格局学说与多顶级学说并无本质的区别,只是后者特别强调了群落变化的连续性。其实连续和非连续也是辩证统一的,它们都是植被格局的真实反映。
2.演替的轨迹
Frelich(2002)提出了关于植被演替轨迹的五种模式:1)平行演替、2)周期性演替、3)趋异演替、4)趋同演替和5)个性化演替(图4)。
平行演替指群落A和群落B在扰动后又各自回复到原来状态的演替。如在北美的北方森林中存在相邻的短叶松和黑云杉林分,在这样的区域发生林分替换火,火前林分是唯一种源,火后植被也维持同样的树木组成(Dix & Swan 1971, Johnson 1992)。
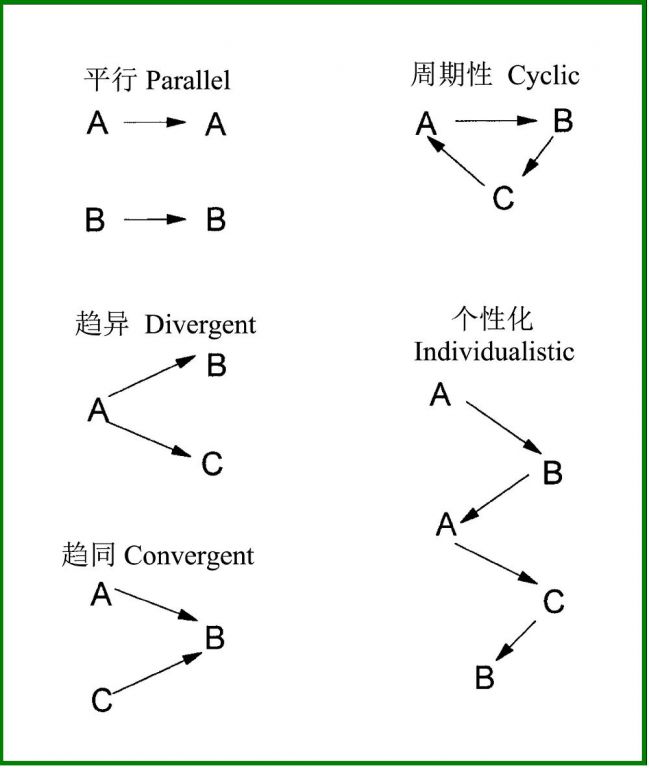
图4 演替方向的五种模式(引自Frelich 2002)
周期性演替主要指由于环境因子的周期性干扰而引起的演替,这种周期性干扰可以是较短的时间尺度(如昆虫的周期性暴发),也可以长久的地质时间尺度(如气候周期)。如在严重的火烧之后,首先山杨侵入,接着红棕及其它中度耐荫种跟进,然后是耐荫性的北方硬木树种和铁杉,一直持续到下一个火烧的来临,又重新回到山杨开始的演替周期(Lorimer 1977,Frelich2002)。
趋同演替就是Clements(1936)提出的经典演替模式,即处于二个或多个状态的植被(状态A和B)随着时间的推移最终趋向于状态C。譬如,二个分别被演替早期或中期树种(如山杨和白松)占据优势的相邻的火后林分均会向耐荫的糖枫和铁杉演替。
趋异演替指一个群落(状态A)随着时间的推移分歧到二个或多个状态(B、C等)的演替,主要是最初的微小差异被逐步放大,最终稳定下来(Wilson and Agnew 1992)。譬如,火后的山杨林分很容易在相邻的三个地点演替成松树、橡树和枫树,这可能由于适合于不同种类的土壤的差异或种源的差异造成的。
五、演替的方向和轨迹依赖于时空尺度
对陆地植物来说,没有什么比气候对其地理格局的影响更为重要的了,气候在不同时间尺度上的波动深刻地影响着植被的变化、演替和演化。谈论演替离不开时空尺度,演替的单顶级和多顶级学说的正确与否其实与时空尺度密切相关。
1.中短时间尺度的植被演替—趋于区域气候顶级
如果以较大(区域)的空间尺度来看,在中等时间尺度(数百年内)的次生演替可能符合气候顶级学说。在地理尺度,气候类型几乎没有争议地决定了大的植被类型,譬如,典型的草地位于降雨量既不能高到孕育森林生态系统的水平,也不可能低到出现沙漠的地步,换言之,在低温、少雨的北方草原地带,决不会演替出热带雨林,而同样在高温的热带地区,多雨的热带雨林地区决不会演替出干旱的刺林植被!因此,从一定气候背景下的植物群系的地理格局来看,演替是有方向性的。这就是说,某一地区的气候类型(如降雨量水平)也基本决定了与此相适应的植被演替的群系类型。
如果以较小(局域)的空间尺度来看,局地非生物环境的异质性、异常复杂的生物间相互作用以及植被地理演化历史等的差异可以呈现丰富多样的植被类型,特别是在群落类型丰富的地域(如湿润的热带-亚热带地区),即可能遵循所谓多顶级学说。
2.长时间地史尺度的植被演替—难觅严格的周期性更替
在较小的时空尺度,你能观察到演替的多种轨迹,如平行、趋同、趋异、个性化和周期性等。但当你把时间尺度扩展到1万年左右,你从孢粉中居然还看不到森林演替的1个周期。
地球上的气温呈现周期性的波动,但无论植物还是动物,都看不到周期性的群落类型。气候在周期性波动中推动物种的革新,旧的物种不断消亡,新的物种不断形成,总体上说,物种的形成速率远高于物种的消亡速率,地球上的生命系统似乎始终处于发展与进化之中。因此,从万年以上的时间尺度来看,似乎不存在所谓的气候顶级,也不存在严格意义上的群落的周期性更替。
结语
有关植被演替的论文或专著可谓汗牛充栋,各种模型之复杂程度令人一筹莫展!人们在纷繁复杂的表象世界中迷失了自我……人们经常争议演替有无方向性、演替的轨迹是否可预测,其实,离开了时空尺度,这些争议将失去意义。过去关于演替的理论或学说如此纷杂,其实原理却十分简单:中短时间尺度的植被演替趋于区域气候顶级,而在长时期的地史尺度上则难觅严格的周期性更替。随着时空尺度的扩展,演替过程呈现出从决定性与可逆性向随机性与不可逆性演进的趋势。这种地史尺度上植被演替的不可逆性恰好与绝大多数物种难逃灭绝厄运的化石证据十分吻合。
参考:从生态学透视生命系统的设计、运作与演化—生态、遗传和进化通过生殖的融合(谢平著,科学出版社,2013,电子版下载:http://wetland.ihb.cas.cn/lwycbw/qt/)
联系方式:沈宏 徐军 027-86780056 Email:wetland@ihb.ac.cn
地址:武汉市武昌区东湖南路7号
访问数量:
